
Проблемы Эволюции |
М.: Наука, 1980
<![if !supportEmptyParas]> <![endif]>
{74}
Глава II
ГЕНЕТИЧЕСКИЕ ОСНОВЫ ПРЕОБРАЗОВАНИЯ ПОПУЛЯЦИЙ
Первый шаг эволюционного процесса — возникновение внутри вида биологически специфичных популяций, особенности которых наследственно закреплены. Отсюда следует, что любая эволюционная теория должна основываться на современных представлениях о законах генетического преобразования популяций. Это делает необходимым предпослать анализу экологических механизмов эволюционного процесса краткий очерк основных выводов популяционной генетики.
Сущность популяционной генетики сводится к изучению законов динамики генетических вариаций (генов и генотипов) в природных и экспериментальных популяциях животных, растений и микроорганизмов. С общебиологической точки зрения эта задача не менее важна, чем центральная проблема общей генетики — изучение структуры наследственных единиц (генов) и механизмов их действия. Для того чтобы понять все возрастающую роль этой молодой науки в развитии эволюционного учения, необходимо обратиться к истории.
Рождение современной генетики, связанное с вторичным открытием законов Менделя, отнюдь не было встречено дарвинистами с восторгом. Это понятно. Ведь суть дарвинистской теории заключается в творческой роли естественного отбора. Между тем генетики утверждали, что единственная известная форма наследственной изменчивости — это мутации. Но возникновение мутаций в отборе не нуждается. Они появляются спонтанно, в готовом виде, и, как думали ранние генетики, особенности мутаций выражены достаточно резко. Максимум, что может сделать отбор,— это отсечь явно неблагоприятные мутации. Роль сита — не очень-то почетная роль для ведущего механизма эволюционного процесса. Этим и определялась реакция дарвинистов. Они не почувствовали в генетике могучего союзника и отрицали роль мутаций как источника изменчивости живых организмов, на основе чего естественный отбор создал все многообразие живых организмов. Распространению этой точки зрения способствовало и то немаловажное обстоятельство, что почти все известные в то время мутации оказались вредными, а в теоретических работах генетиков того периода проявились механистические тенденции. С другой стороны, дарвинисты еще не понимали, что закон расщепления, независимого распределения признаков и относительного постоянства единиц наследственности (генов) снимает ряд трудностей теории Дарвина.
{75}
Само собой понятно, что в трудах разных ученых тенденция противопоставления дарвинизма и менделизма проявлялась в разной степени и в разных формах, но в целом почти 30-летний период начала XX в. можно рассматривать как период самостоятельного развития эволюционного учения и генетики. Решающий шаг к взаимному обогащению этих ведущих разделов естествознания сделал в 1926 г. С. С. Четвериков в статье «О некоторых моментах эволюционного процесса с точки зрения современной генетики». В нашу задачу не входит анализ этой замечательной статьи, положившей начало новому направлению и в генетике, и в эволюционном учении. Отметим лишь главное. С. С. Четвериков раскрыл роль дискретных естественных единиц в изменчивости популяции и показал, что на основе мутаций в природных популяциях животных накапливается огромный потенциал изменчивости — «скрытая изменчивость». Этот наследственный потенциал, механизм формирования которого вполне соответствует законам генетики, и есть тот материал, на основе которого работает естественный отбор. Основной вывод С. С. Четверикова получил подтверждение и развитие в трудах плеяды замечательных генетиков, среди которых видное место занимают русские исследователи (Н. П. Дубинин, Н. В. Тимофеев-Ресовский и др.). История науки еще сделает предметом специального анализа этот интереснейший период в развитии биологии. За каких-нибудь десяток лет генетика, рассматривающаяся до работ С. С. Четверикова неясным спутником дарвинизма, стала его основой, а эволюционные работы, не учитывающие достижения генетики, вдруг стали простым анахронизмом. Стало ясным, что именно мутации и есть тот материал, на котором основывается эволюция, а следовательно, все закономерности, касающиеся цитологических основ наследственности и фенотипического проявления мутаций, имеют прямое отношение к дарвинизму, к теории естественного отбора. Симптоматично само название одной из важнейших работ этого периода: «Генетическая теория естественного отбора» [Fisher, 1930]. Возникла новая система идей, суть которой сводится к следующему, источник изменчивости — мутации генов и их рекомбинации. Особо подчеркивается исключительная роль рекомбинаций. Если наследственность животного определяется 1000 генами, каждый из которых может проявляться в 10 аллелях, то это значит, что на основе рекомбинаций может возникнуть 101000 генотипов. Это число превышает число электронов в видимой части Вселенной.
Что же определяет распространение в популяции отдельных генов, что определяет их сочетание в определенные генотипы, наилучшим образом приспособленные к условиям внешней среды? Какую роль в этом процессе играет мутирование? Ответить на эти вопросы методами классической генетики оказалось невозможным. Возникло новое направление генетики — популяционная генетика.
Популяция с точки зрения генетика — элементарная совокупность особей вида, в пределах которой реализуется свободное скре-
{76}
щивание, осуществляется свободный обмен генами. Это определяет генетическое своеобразие популяции, ее отличия от других популяций вида. Однако генетическое единство популяции всегда сочетается с ее генетическим разнообразием 1. Даже родные братья и сестры генетически не идентичны. Это особенно ярко было показано на плодовитых животных, например амфибиях. Потомство от одной пары животных представляет собой популяцию в миниатюре, генетическая разнородность которой остается значительной. Отсюда следует, что любое скрещивание приводит к возникновению новых генетических вариантов.
Это положение существенно отличается от представления генетиков начала века. Не крупные, бросающиеся в глаза наследственные изменения (макромутации — коротконогость овец, резкие отличия в окраске птиц или млекопитающих, изменение структуры волоса или пера и т. д.) определяют различия между особями популяции. Макромутации, как правило, вредны и в природных популяциях проявляются относительно редко. Генетическая индивидуальность животных детерминируется мелкими, нейтральными мутациями, ничтожными изменениями наследственного аппарата клетки, определяющими такие ничтожные особенности животных, как чуть-чуть более светлую или темную окраску, чуть-чуть большие или меньшие размеры тела, ничтожные различия в уровне метаболизма и т. п. Изучая внешние проявления подобных малых мутаций, генетики пришли к выводам, имеющим первостепенное значение для познания механизмов эволюционного процесса. Оказалось, что громадное большинство признаков, которыми отличаются животные в пределах популяции, определяется не одним, а комплексом генов (полигенная детерминированность признаков) и еще большим числом генов-модификаторов. Это страхует организм от случайного вредного изменения генотипа. Если данная особенность организма определяется молекулярной структурой одного из тысяч локусов хромосом, то изменение этого локуса (в результате мутаций или рекомбинаций при скрещивании) неизбежно вызовет заметное изменение морфофизиологических особенностей животного. Если же признак определяется структурой многих локусов, тем более расположенных в разных хромосомах то возможности случайного нарушения нормального развития сводятся к минимуму. Поэтому понятно, что наиболее важные признаки организма (в особенности его физиологические особенности) застрахованы указанным путем особенно надежно 2. Свойства части (гена) определяются свойством целого (генома, а весьма вероятно — и половой клетки в целом). Появляются данные,
1 Экспериментально доказано, что даже высокое давление отбора не приводит к генетической однородности популяции. Генетическое разнообразие в модельной популяции дрозофилы сохранилось и после 70 поколений, подвергавшихся отбору [Stebbins, 1965].
2 С особой убедительностью это было показано в посмертной книге И. И. Шмальгаузена [1964].
{77}
показывающие, что дополнительная наследственная информация может кодироваться на более высоком (по сравнению с триплетами ДНК) структурном уровне [Sand, 1965]. Таким путем современная генетика преодолевает известный механицизм ранних представителей. Эти открытия имеют важные следствия в популяционной генетике. Так как малые мутации в большинстве случаев не оказывают видимого влияния на жизнеспособность животного, то это создает условия для их накопления в общем генофонде популяции. В популяции возникает громадный резерв генетических потенций.
Другое, не менее существенное открытие популяционной генетики заключается в повышенной жизнеспособности гетерозигот. Сочетание в едином генотипе разных аллельных генов (разных генетических потенций, унаследованных от отца и матери) повышает жизнеспособность организма. Поэтому даже в том случае, если мутация оказывается вредной, она не исчезает из популяции, так как повышенная гибель гомозигот уравновешивается снижением смертности гетерозигот. Это приводит к тому, что; например, в популяциях человека сохраняются гены, вызывающие в гомозиготном состоянии тяжелые заболевания (в целом их присутствие в популяции оказывается полезным). Естественно, что и это имеет следствием непрерывное обогащение генофонда популяции. Ценность индивида определяется вкладом в популяцию [Stebbins, 1965].
Специальный аспект этой проблемы, представляющий для экологии особый интерес, отмечается в исследованиях Левонтина [Lewontin, 1955] с чистыми и смешанными культурами дрозофилы. Оказалось, что жизнеспособность отдельных генотипов зависит от генетической структуры популяции в целом и не может быть определена на основе изучения жизнеспособности чистых культур отдельных генотипов. В этом отношении исключительный интерес представляют исследования по генетике мимикрии. Изучение различных видов бабочек показало, что подражательная окраска обычно детерминирована одним доминантным геном, но совершенство мимикрии зависит от генома в целом [Sheppard, 1965]. Изучая полиморфный вид Papilio dardanus, Форд [Ford, 1963] установил, что в результате мутаций возникает лишь приблизительное сходство подражателя с моделью, а его совершенствование — это результат преобразования генотипа. Если основной ген ввести в генотип другой расы, то совершенство мимикрии снижается. Снижается и степень доминирования основного гена. Это свидетельствует о том, что и сама доминантность вырабатывалась отбором путем преобразования генотипа разных внутривидовых форм (автор полагает, что это связано с действием супергенов).
Третий вывод рассматриваемого направления популяционной генетики имеет, пожалуй, для развития эволюционного учения еще большее значение. Коль скоро важнейшие биологические особенности организма определяются не единичными генами, а их комплексами и поскольку вредные (в том числе и летальные)
{78}
мутации могут оказаться полезными в гетерозиготном состоянии то из этого следует, что ценность любой мутации определяется не ее индивидуальными свойствами, а свойствами генотипа в целом. Но так как скрещивание приводит к постоянному перемешиванию генов и постоянному изменению генотипов, то в конечном итоге ценность отдельных мутаций определяется свойствами общего генофонда популяции в целом. Весьма существенно, что уже довольно давно были получены прямые экспериментальные доказательства, что гетерозиготы обладают совершенными гомеостатическими реакциями, повышающими приспособленность животных к изменению условий среды [Dobzhansky, Levine, 1955].
Этот вывод имеет исключительное значение. Он показывает, что популяция — единая целостная система: изменение отдельных генотипов влияет на общий генофонд популяции, но и изменение генофонда изменяет роль отдельных генотипов и даже отдельных генов в развитии системы. Отсюда единственно возможное заключение: элементарной единицей эволюционного процесса являются не отдельные особи, а популяции. Это положение стало основой современного эволюционного учения, его справедливость ежегодно подтверждается новыми экспериментальными исследованиями.
С другой стороны, сказанное ранее показывает, почему, несмотря на то что единичные наследственные изменения (мутации) по своей природе дискретны, изменчивость организмов имеет непрерывный характер. Эта непрерывность есть следствие совокупного действия множества генов и их модификаторов.
Распространение генетических представлений с уровня индивидов на уровень популяционный заставило биологов больше внимания уделить стохастическим процессам и с исторической неизбежностью вызвало появление цикла работ, который может быть назван математической теорией преобразования популяций [Fisher, 1930; Haldane, 1954, 1957; Wright, 1948, 1955]. Эта теория впервые использовала математическое моделирование как средство анализа эволюционных процессов и выявила наиболее общие закономерности, определяющие относительную роль давления отбора, размеров популяции, степени изоляции, типа динамики численности в изменении генетической структуры популяции. Сущность математического направления в популяционной генетике удачно сформулирована Райтом. Он указывает, что математическая теория связывает в единую систему генетику особей и генетику популяций; она основана на моделировании структуры популяций, которая подвергается корректировке на основе сопоставления моделей с полевыми наблюдениями. Важнейшим итогом этого направления работ явилось предсказание явления так называемого дрейфа генов (genetic drift), анализ которого привел к учению о генетико-автоматических процессах 1. Исследованию степени соответствия
1 Основы этого учения были по существу заложены С. С. Четвериковым в его представлениях о роли «воли жизни» в микроэволюционном процессе.
{79}
математических моделей явлениям, протекающим в природе, был посвящен ряд специальных работ [Wright, 1955; Dobzhansky, 1954, 1955, 1958; Lerner, 1965; Falconer, 1965] и симпозиумов в Италии (1953 г.), США (1960 г.) и в других странах. Тем не менее, как мы попытаемся показать в последующих главах, разрыв между моделью и реальностью все еще продолжает оставаться значительным, что породило некоторые ошибочные представления, имеющие большое значение для развития эволюционной теории.
Необходимость приблизить математическое моделирование эволюционных преобразований к реальной природной обстановке ощущалась уже давно. Это удачно сформулировал Вильямсон [Williamson, 1957], указавший, что математическая теория внутривидовых отношений исходит из упрощенных представлений об однородности особей одного вида и об однородности и замкнутости среды их обитания. Шеппард [Sheppard, 1965] как о чем-то хорошо известном пишет, что современные исследования показали, что отбор работает с большей эффективностью, чем это следует из математического моделирования.
С другой стороны, известные японские генетики Кимура и Ота свою программную статью [Kimura, Ohta, 1974] предваряют утверждением, что эволюция на молекулярном (генном) уровне в значительно большей степени определяется мутационным давлением и случайным дрейфом генов, чем это представляется ортодоксам современного неодарвинизма. В доказательство они ссылаются на работы, показывающие, что скорость эволюции, выраженная числом аминокислотных замещений в различных протеинах, практически тождественна в разных филогенетических линиях и сохраняется постоянной до тех пор, пока сохраняется неизменной функция белка или не изменится его третичная структура.
В более ранней работе Кимура показал, что число аминокислотных различий между a- и b-гемоглобином человека примерно равно числу различий между a-цепью карпа и b-гемоглобином человека [цит. по Kimura, Ohta, 1974]. Это значит, что мутации генных локусов, кодирующих a- и b-цепь, происходили с одинаковой скоростью в двух эволюционных линиях, разошедшихся около 500 млн. лет тому назад. Авторы полагают, что эти и многие аналогичные им данные свидетельствуют о постоянстве процесса молекулярной эволюции.
Примерное постоянство скорости эволюционных преобразований других белков (цитохром С, альбумин фибринопептиды и др.) было продемонстрировано различными авторами. Есть и данные, противоречащие этому утверждению (см., например, [Langley, Pitch, 1973]), однако если сравнение проведено за достаточно длительный период времени, то тезис о постоянстве эволюции белковых молекул кажется в настоящее время хорошо обоснованным. Объяснить это постоянство с позиций неодарвинизма Кимура и Ота [Kimura, Ohta, 1974] считают практически невозможным.
{80}
Метод исследования эволюции, основанный на изучении фенотипов, кажется им ограниченным.
Исследования, проведенные в рассматриваемом направлении показали, что функционально менее важные молекулы или части молекул изменяются в процессе филогенеза быстрее, чем функционально существенные. Так, и в a- и в b-цепи гемоглобина изменение поверхности частей молекулы происходит в 10 раз быстрее чем в молекулярной структуре гена. Связанные с геном гистидины оказались абсолютно неизменными в течение всей эволюции позвоночных (около 500 млн. лет). Кимура и Ота полагают, что скорость эволюционных субституций в молекулах, изменение которых легко может быть «принято» организмом, близка к скорости спонтанного мутирования, так как изменения молекул легко могут вписаться в популяции, подчиняясь закономерностям дрейфа генов (в противном случае, согласно авторам, пришлось бы предполагать действие интенсивного положительного отбора мутаций, не имеющего видимого биологического значения).
Мутантные субституции, в меньшей степени нарушающие структуру и функции молекулы (консервативные субституции), встречаются чаще. Это соответствует представлению Фича [Fitch W. М., 1972] о «сопутствующе изменяющихся кодонах» (concomitantly variable codons) — коварионах (covarious). Только 10% кодонов в цитохроме С могут принять мутацию в любой момент эволюции. Фич установил, что в a-гемоглобине около 35% коварионов, а в фибринопептиде А — 100%. Из его исследований следует, что если число аминокислотных субституций вычислять на основе коварионов, то скорость эволюции цитохрома С, a-гемоглобина и фибринопептида А оказывается одинаковой.
Дупликация генов считается обязательным условием возникновения гена, обладающего новыми функциями. Наличие генов в двух копиях создает предпосылки для накопления мутаций в одной из них. Это особенно ясно было показано Оно [1973].
Кимура и Ота утверждают, что элиминация явно вредных мутаций и случайное закрепление нейтральных или почти нейтральных мутаций происходят в процессе эволюции значительно чаще, чем это представляется неодарвинистам. Это утверждение авторы рассматривают как развитие своих более ранних представлений о важной роли случайных мутаций в молекулярной эволюции: давление мутаций ведет к эволюционным изменениям, как только преодолевается барьер отрицательного отбора.
Рассмотренное направление популяционной генетики исследует законы формирования генофонда популяции. Другое направление изучает генетическое преобразование популяции при изменении условий внешней среды (изменении направления отбора). В соответствии с изменением направления отбора происходит изменение частоты встречаемости разных генотипов и изменение средней нормы изменчивости популяции в целом. Этот процесс может быть легче всего понят на конкретном примере. Стрептоми-
{81}
цин в концентрации 25 мг/кг останавливает рост кишечной бактерии Escherichia coli. Однако если несколько миллионов бактерий выращивать на питательной среде, содержащей стрептомицин, то наблюдатель вскоре обнаружит, что через несколько поколений пост Е. coli возобновляется и не прекращается даже при высоких концентрациях антибиотика. Специальный анализ показал, что среди миллионов бактерий (напомним, что популяции бактерий, подобно популяциям любых других существ, генетически разнородны) оказались и такие, которые обладают наследственной стойкостью к стрептомицину. Естественно, что они не были уничтожены и дали новые поколения, не восприимчивые к стрептомицину.
Точно таким же путем в условиях эксперимента были созданы ядостойкие популяции различных насекомых. Они возникают и в природе при бездумном использовании стандартных ядохимикатов. В районах, регулярно обрабатываемых ДДТ, устойчивость некоторых насекомых к яду за 10 лет увеличилась в 100 раз. Резко возросла и устойчивость к ДДТ лягушек [Ferguson, 1963]. Важно отметить, что устойчивость насекомых к ДДТ определяется генами, расположенными во многих хромосомах [Crow, 1960].
Экспериментальные исследования этого направления к настоящему времени насчитываются многими сотнями (общее число работ по популяционной генетике уже давно перевалило за 2000). Их главный итог сводится к нескольким положениям, столь твердо доказанным, что они заслуживали бы возведения в ранг законов популяционной биологии.
1. Чем больше генетическая разнородность популяции и чем больше ее генофонд, тем выше ее жизнеспособность, тем выше ее экологическая способность, тем быстрее и полнее она преобразуется под влиянием измененной среды и соответственного изменения направления отбора. Экспериментально показано, что отбор в течение 15 поколений изменяет устойчивость некоторых экспериментальных популяций Drosophila melanagaster к ДДТ в 600 раз [Benett, 1960].
2. Доказано, что в отдельных случаях преобразование популяции вызывается изменением частоты распространения моногенно детермированных признаков 1. В других, значительно более частых
1 Блестящий пример изменения генетической структуры популяции подобного типа — возникновение так называемого индустриального меланизма Biston betularia [Kettlewell, 1956] в Англии. Еще в начале XIX в. светлая форма этой бабочки (cc), окраска которой гармонировала с цветом светлых лишайников на коре берез, была широко распространена. Однако по мере того, как под влиянием промышленных дымов кора берез стала темнеть, преимущество стал получать темный мутант carbonaria (СС). В настоящее время cc стали редкостью. Было установлено, что светлая форма действительно чаще поедается птицами, чем покровительственно окрашенные СС и сС.
В отдельных (вероятно, редких) случаях моногенно детерминированными оказываются и биохимические признаки. Это было показано, в частности, при анализе межлинейных различий мышей по биосинтезу различных кортикостероидов [Badr, Spickett, 1965].
{82}
случаях изменение отбора ведет к изменению сочетания производителей, к изменению селекционной ценности различного сочетания совокупно действующих генов. Таким путем на основе исходного генофонда возникают новые генотипы, отсутствующие в исходных популяциях.
3. Отбор в течение многих поколений (громадное большинство природных популяций существует сотни и тысячи лет) создал наилучшим образом сбалансированные генотипы и наилучшим образом сбалансированный популяционный генофонд. В этой сбалансированной системе жизнеспособность отдельных генотипов определяется комплексом других. Это значит, что даже если ни темп мутирования, ни его характер не изменяются (зависимость характера мутирования от свойств генотипов — один из сложнейших и еще не решенных вопросов популяционной генетики 1), то и в этом случае в измененной в результате отбора популяции новые мутации будут иметь уже новое значение, так как они служат основанием для формирования новых генотипов. Это обеспечивает принципиальную безграничность эволюционного прогресса.
Мы видим, что выводы популяционной генетики не противоречат гениальным принципам классического дарвинизма. Наоборот, они наполняют конкретным содержанием расплывчатое понятие Дарвина «неопределенная изменчивость» и позволяют вскрыть конкретные механизмы начальных стадий эволюционного процесса. Однако популяционная генетика породила и некоторые теоретические трудности. Эти трудности прямо или косвенно оказались связанными с анализом темпов эволюционного процесса.
Математический анализ возможной эффективности индивидуального отбора, проведенный Холденом [Haldane, 1954], привел автора к выводу, что в случае горотелической эволюции субституция одного аллеля требует около 300 поколений. Холден полагает, что его вывод соответствует имеющимся данным, характеризующим «средний» темп эволюционного процесса. В это же время принципиально сходные выводы были сделаны на основании экспериментального изучения изменений полигенно детерминированных признаков у дрозофилы [Buzzati-Traverso, 1955]. Было показано, что под влиянием отбора в модельных популяциях такие признаки, как размеры тела или длина крыла, изменяются со скоростью 0,00024% за поколение. Это примерно соответствует максимальной скорости филогенетических преобразований, установленных палеонтологами (индекс высоты черепа в эволюции человека). Автор делает далеко идущий вывод о том, что известные генетические процессы (воспроизводимые экспериментально)
1 В опытах С. М. Гершензона [1965] о наездником Marmoniella vitnpennis были получены данные, показывающие, что у самцов, происходящих от скрещивания двух линий разного географического происхождения, частота возникновения доминантных мутаций повышена.
{83}
адекватны механизмам филогенетических преобразований, которые фиксирует палеонтолог. Несмотря на то что в этой главе мы стремимся свести к минимуму элемент дискуссии, нельзя не отметить, что оба автора, выражающие точку зрения большой группы исследователей, рассматривают эволюцию как процесс горотелический. Между тем нет ровно никаких оснований считать доказанным, что скорость эволюционных преобразований стабильна. Однако даже если рассматривать процесс эволюции как горотелический, то и при этом ряд фактов заставляет признать наличие еще каких-то механизмов преобразования популяций, сводимых к понятию «естественный отбор» лишь при самой широкой его трактовке. Ограничимся немногими примерами, так как специальный анализ этого вопроса заслуживает особого рассмотрения.
Завезенный в Америку воробей за 100 лет изменился в такой степени, что по всем таксономическим правилам его следовало бы возвести в ранг резко выраженного подвида. Учитывая различия в условиях существования воробья на новой и старой родине, подобная скорость формообразования явно не укладывается в рамки теории.
О большой скорости эволюционных преобразований свидетельствуют и другие факторы. Озеро Ланао на о-ве Минданао (Филиппины) существует не более 10000 лет. За это время здесь образовалось не менее 18 видов и 4 родов рыб [Myers, 1960]. Видообразование среди рыб в некоторых озерах Северной Америки происходило столь же быстро [Hubbs, Raney, 1946; Miller, 1961]. В озере Набугато (Уганда) не менее 5 видов рыб (Cichlidae) сформировались за 4000 лет. Домовая мышь, завезенная на Фарерские острова около 300 лет тому назад, изменилась настолько, что некоторые авторы считают ее новым видом [Мауг, 1963]. Весьма вероятно, что подобные явления отнюдь не уникальны, мы просто не научились их наблюдать. Слишком мало внимания уделяется исследованиям морфофизиологических особенностей акклиматизированных форм [Шварц, 1963; Шапошников, 1958], а цитогенетические исследования в широком географическом плане еще до сих пор остаются редкими. Возможно поэтому целый ряд микроэволюционных явлений, протекающих перед нашими глазами, остается незамеченным. Об этом свидетельствуют наблюдения, показавшие, что с 1940 по 1957 г. генетическая структура (частота встречаемости различных хромосомных перестроек) Drosophila pseudoobscura существенно изменялась на громадной территории юго-востока США [Dobzhansky, 1958]. Противоположный пример может быть заимствован из хорошо изученной палеонтологической истории лошадей. У ископаемых предков нашей лошади в связи с приспособлением к питанию жесткой пищей происходило неуклонное увеличение диаметра коренных зубов. Но происходило оно со средней скоростью 0,2 мм за миллион лет. При этом диапазон изменчивости в пределах отдельных популяций достигает 3 мм! Сопоставление этих примеров показывает, что должны быть, помимо
{84}
давления естественного отбора, еще какие-то факторы, определяющие скорость эволюционных преобразований популяций. Гипотезы рассматривающие эти факторы, уже более 20 лет тому назад оформились в теорию генетико-автоматических процессов. Ее развитие связано с именами С. С. Четверикова, С. Райта, Н. П. Дубинина, Э. Майра и некоторых других генетиков и зоологов.
Сущность представлений о генетико-автоматических процессах может быть сведена к следующему. Если какой-то изолированный участок пространства (остров для наземных животных, озеро для рыб и т. п.) заселяется представителями определенного вида, то поселенцы не являются полноценными представителями исходной популяции. Генофонд новой популяции не только обеднен, но и специфичен, так как определяется генотипом особей-основателей. Поскольку заселение новых территорий или акваторий происходит случайно, то и генетическая структура новой популяции в значительной степени обязана случайности. Само собой понятно, что и на новой родине популяция будет подвергаться силам отбора, но так как отбор всегда работает на основе наличного генофонда, то и на результат отбора исходный состав новой популяции не может не повлиять, тем более что приток новых особей извне прекращен (изоляция!). Быстро возникает новая популяция, изменяющаяся в своеобразном направлении. Так, например, на архипелаге островов возникает группа близких форм (подвидов, видов), обособление и морфологическая дифференциация которых происходит значительно быстрее, чем на сплошном участке ареала. Особенности этих форм трудно объяснить только действием естественного отбора, но они могут быть объяснены теорией генетико-автоматических процессов.
Легко понять, что вполне аналогичный принцип действует не только в пространстве, но и во времени. После резкого спада численности вида, вызванного неспецифическим фактором среды (наводнения, бури, весенние снегопады и т. п.), популяция восстанавливается за счет немногих оставшихся в живых особей. Их генофонд по понятным причинам не совпадает с исходным генофондом популяции, поэтому при восстановлении численности срабатывает тот же самый механизм случайного основателя, что и при пространственной изоляции. Не вдаваясь в детали, полезно указать, что согласно большинству теоретических представлении эволюционные преобразования популяций происходят особенно быстро в тех случаях, когда вид представлен относительно изолированными популяциями средних размеров.
В новейшее время связь между динамикой численности и изменением генетического состава популяции экспериментально изучена Кребсом с сотрудниками [Krebs et al., 1973; Krebs, Myers. 1974; Myers, Krebs, 1974]. Заслуживают внимания не только результаты экспериментов, но и логика исследований. Отмечено, что 3–4-летний цикл численности наблюдается у ряда видов грызунов Северной Америки и Европы (отсутствует в тропиках и в
{85}
южном полушарии). Авторы обращают внимание, что весенний крах популяции сильнее выражен у самцов, чем у самок, и что после «завершения цикла» судьбу популяции трудно предсказать. Новые подходы к изучению динамики численности грызунов Кребс датирует 1950 годом, выходом в свет первых работ Христиана (Christian), объяснявших крах популяции стресс-реакцией при высокой численности. Однако исследования Читти [Chitti, 1960] доказали, что ожидаемый эффект высокой численности наблюдается не у животных, развивающихся при повышенной плотности, а у их потомков. Читти предположил, что в условиях напряженных контактов между отдельными особями способность к размножению сохраняют генетически своеобразные животные. Это утверждение явилось исходным в работах группы Кребса, которые изучили изменения в поведении животных (в частности, их агрессивности) на разных фазах цикла численности. На Microtus pennsylvanicus и М. ochrogaster (изучено 1140 самцов каждого вида) было показано, что самцы в период пика численности отличаются повышенной агрессивностью. Для того чтобы установить, связано ли изменение в поведении животных с изменением генетического состава популяции, были изучены электрофореграммы крови. Вероятные отличия в аминокислотном составе белков плазмы авторы рассматривают в качестве «случайных маркеров, благодаря которым мы можем наблюдать генетические изменения в популяции» [Myers, Krebs, 1974]. Авторы работали с двумя протеиновыми системами, трансферрином (Tf-белок, транспортирующий железо) и лейцин-аминопептидазой (LAP). Оба белка представлены в популяциях грызунов в двух вариантах. Было показано, что изменение численности действительно сопровождается изменением в, частоте сравниваемых генов. Особо резкие изменения наблюдались в фазе падения численности. Направление изменений в независимых циклах совпадало. ТfC/ТfC выживают в большем числе во время нарастания численности, TfC/TfE — в период пика, TfE/TfE— в период падения. Отмечены различия между резидентами и мигрантами. В период нарастания численности выживаемость высокая, селекционные преимущества имеют быстро размножающиеся генотипы 1. Оставалось, однако, не ясно, является ли изменение генетического состава популяции следствием или причиной изменений численности (как полагал Читти). Авторы ставили эксперименты в вольерах. Было установлено, что в этих условиях плотность в 20 раз превышает плотность свободных популяций, но сильно поврежденная растительность полностью восстанавливается весной. Становится очевидным, полагают авторы, что стресс, связанный с повышением плотности, не может сам по себе, вызвать крах популяции. Затем авторы в течение двух лет изы-
1 Явная логическая ошибка. Наоборот, в период, когда селекционные преимущества «специальных» генотипов не сказываются, в большом числе размножаются генотипы, отличающиеся скоростью размножения.
{86}
мали часть популяции и. сравнивали резидентов с мигрантами. Среди мигрантов преобладали молодые самки. Таким образом репродуктивный потенциал колонистов существенно повышен. При этом генотип самок-вселенцев резко отличался от генотипа резидентов. Следовательно, генофонд популяции при высокой и низкой численности различен. Авторы полагают, что в популяции существуют два «типа» генотипов — способных быстро размножаться и способных жить при высокой численности. Причину падения численности популяции следует искать в ее генетической структуре.
Проявление генетико-автоматических процессов в пространстве получило известность как принцип Майра, во времени — как принцип Райта. В ряде работ они достаточно четко формулируют теоретическое кредо «синтетической теории эволюции».
Райт [Wright, 1948, 1955, 1959] подчеркивает, что эволюция определяется действием ряда факторов: мутаций, отбора, случайных факторов. Наиболее эффективным «механизмом эволюции» Райт считает подразделение видовой популяции на частично изолированные локальные популяции, дифференциация которых происходит под совместным действием случайных процессов (принцип основателя) и индивидуального и межпопуляционного (interdeme selection) отбора. Еще более определенен в своих суждениях Майр [Mayr, 1954]. «Изоляция немногих индивидов («основателей») из изменчивой популяции, расположенной по пути потока генов, медленно протекающего по ареалу любого вида, производит внезапное изменение генетической среды большинства локусов. Это изменение — действительно наиболее сильное генетическое изменение (за исключением полиплоидии и гибридизации), которое может возникнуть в популяции, так как оно может воздействовать на все локусы одновременно. Действительно, оно может принять характер «генетической революции». В дальнейшем эта «генетическая революция», возникающая благодаря изоляции популяции-основателя, может приобрести свойства цепной реакции. Изменение в любом локусе в свою очередь воздействует на селективную ценность во многих других локусах, пока система не достигнет нового состояния равновесия». В одной из своих последних работ Майр [Mayr, 1965] находит возможным утверждать, что под влиянием отбора генотипы почти автоматически преобразуются гармоническим образом.
Таким образом, генетики, а следом за ними и многие зоологи и ботаники пришли к выводу, что эволюционный процесс определяется естественным отбором, изоляцией и динамикой численности (волны жизни). Разные авторы придают этим факторам эволюционного процесса различные значения, но в целом эта концепция признается большинством биологов. Как уже указывалось, наиболее крупные эволюционисты отлично понимают, что в своем современном виде теория эволюции еще далека от абсолютного совершенства. Но ведь это можно сказать про любую естественно-
{87}
научную теорию. И, подобно любой теории, она должна развиваться. Каковы же пути развития теории эволюции органического мира? Здесь мы покидаем твердую почву уже давно завоеванных наукой фактов и становимся на неизмеримо более зыбкий грунт гипотез.
Несомненно, что в ближайшем будущем значение популяционно-генетических исследований не только не уменьшится, но и значительно возрастет. Об этом свидетельствуют работы последних лет, вскрывшие ряд принципиально новых закономерностей. Так, за последний 10-летний период было показано, что уровень интеграции генотипа в процессе эволюции возрастает, что обогащение популяционного генофонда сопровождается увеличением резерва генетических потенций отдельных индивидов и соответственно с этим роль изоляции зависит от того, из каких популяций происходят животные-основатели 1, что при скрещивании двух популяций, спустя несколько десятков поколений, возникает новая популяция, новая полигенная система, лучше приспособленная к условиям внешней среды. Трудно переоценить значение подобных исследований для совершенствования теории эволюции. Но нам кажется, что любая степень их развития не решит поставленной задачи во всей ее неимоверной сложности. На чем основывается это ответственное утверждение?
Несомненно, первый шаг эволюционного процесса заключается в преобразовании популяций. В этом направлении популяционная генетика сделала исключительно много: основные пути процесса вскрыты и могут быть воспроизведены в эксперименте. Однако эволюция не исчерпывается ее первым шагом. Объяснить эволюцию — это значит вскрыть механизмы видообразования с той степенью детальности, которая позволила бы управлять ими.
1 Как мы попытаемся показать в дальнейшем, это положение имеет исключительное значение для понимания экологических механизмов эволюционного процесса. Важно поэтому подчеркнуть, что оно основано на строгих экспериментальных фактах [Carson, 1961]. Популяции Drosophila robusta из центра ареала вида, отличающиеся высокой степенью генетической разнородности, характеризуются большой генетической емкостью (storage capacity) отдельных кариотипов. Это доказывается тем, что модельные популяции, возникающие от одной пары мух-основателей, не отличались от популяций, основателями которых было большее число индивидов. С другой стороны, было показано, что обогащение генофонда подобных популяций путем скрещивания их с мухами из периферийных популяций не вызывает обычного эффекта повышения жизнеспособности популяции. Это свидетельствует, по мнению автора, о ее генетической насыщенности. Дело генетиков — проанализировать механизмы этих интереснейших явлений. Здесь же важно отметить, что они хорошо согласуются со многими наблюдениями экологов, которые имеют в рамках нашей темы исключительное значение.
{88}
Глава III
ЭКОЛОГИЧЕСКАЯ ОБУСЛОВЛЕННОСТЬ ФЕНОТИПА
Основанное на огромном экспериментальном материале и современных теоретических представлениях убеждение в ненаследуемости благоприобретенных признаков явилось объективной и субъективной причиной резкого снижения интереса к исследованию непосредственных реакций организмов на изменения условий их существования и развития. Между тем характер фенотипических реализации наследственной программы играет решающую роль в эволюционном процессе. Важно поэтому разобраться в том, каков диапазон фенотипической изменчивости в разных группах организмов. Здесь в дальнейшем под фенотипическими изменениями мы понимаем любые морфофизиологические изменения животных, не связанные с изменениями их наследственности.
Убеждение в том, что фенотипическая изменчивость прямого отношения к эволюционному процессу не имеет, приводит к тому, что даже в серьезных современных монографиях ограничиваются хорошо известными примерами, указывающими на связь фенооблика животных и растений с условиями их развития. Вот уже почти сто лет из учебника в учебник, из сводки в сводку кочуют одни и те же примеры, изменение формы листа стрелолиста при росте в воде и под водой, увеличение массы мышц в процессе тренировки и т. и. Важно поэтому показать, что, во-первых, и указанные простейшие примеры различной фенотипической реализации одной и той же генетической программы в действительности отнюдь не простые биологические явления и, во-вторых, они затрагивают лишь одну из многих сторон проблемы, оставляя в тени не менее важные и интересные закономерности.
Простейшая фенотипическая реакция организма: в ответ на повышение физических нагрузок масса мышц возрастает. Но механизм этой реакции весьма сложен. Ограничимся кратким описанием эндокринных механизмов ее реализации.
В ответ на изменение условий среды, требующих интенсификации двигательной активности животного, увеличивается продукция анаболических стероидных гормонов, производных тестостерона, андростерон-диола и 19-нортестостерона. Биологические свойства анаболических гормонов хорошо известны, но механизм их действия изучен хуже. Показано, однако, что они воздействуют на белковый обмен, задерживают азот в клетках, стимулируют синтез белка. Синтез и накопление белка в скелетных мышцах,
{89}
миокарде, почках, печени, нервных клетках увеличиваются. Под влиянием анаболитов изменяется и минеральный обмен организма: в клетках задерживается калий, фосфор и сера в необходимых для синтеза белка соотношениях. Как следствие всех этих процессов происходит значительный рост мышечной массы [Семенов, Шаев, 1975]. Таким образом, фиксируемая простейшая фенотипическая реакция имеет в своей основе изменение активности внутриклеточных ферментных систем, принимающих непосредственное участие в синтезе белка. Биологическая целесообразность комплекса указанных реакций подчеркивается и тем обстоятельством, что анаболические гормоны способны компенсировать метаболические изменения, вызываемые стрессом. В естественной среде обитания ситуации, требующие повышенной двигательной активности, как правило, вызывают состояние психического напряжения и усиленную секрецию гормонов. Однако, с другой стороны, усиленная секреция анаболических гормонов вызывает множество сопутствующих изменений физиологии организма, многие из которых могут привести к нежелательным (возможно, патологическим) последствиям (нарушение генеративного цикла 1, нарушение нормальной функции печени и почек и т. п.).
Становится очевидным, что даже простейшие фенотипические реакции в действительности определяются сложнейшими физиологическими механизмами, совершенство которых проверено отбором в процессе эволюции любого вида.
Раньше (буквально лет 10 назад) дело казалось относительно простым: если популяции отличались «стабильными» признаками — их особенности считались наследственными, в противном случае — ненаследственными, фенотипическими. Сейчас положение существенно изменилось. Было показано, что даже такие «стабильные» признаки, как краниологические особенности млекопитающих, в определенных условиях подвержены сильнейшей фенотипической изменчивости [Dehnel, 1949], а изменения пропорций тела и черепа животных (один из излюбленных признаков таксономистов) могут быть вызваны не менее просто, чем изменения веса резервного жира (пример крайне лабильного показателя). Изменение скорости роста вызывает изменения конституции животных, которые по своим масштабам не уступают различиям между многими подвидами.
Работы школы Денеля указывают на возможность очень существенных межпопуляционных отличий, основанных на фенотипических механизмах. Об этом свидетельствуют и другие исследования, в том числе довольно старые. Так, Клятт [Klatt, 1926, цит. по Hesse-Doflein, 1943] еще в 20-х годах указывал, что кормление тритонов мясом моллюсков приводит к ускорению их роста,
1 Экологически аналогичное явление: потеря репродуктивной функции у крыс, адаптированных к низкому барометрическому давлению.
{90}
которое связано с гиперфункцией гипофиза. При этом меняется и форма тела: голова укорачивается, нижняя челюсть удлиняется. Работа Клятта находит себе подтверждение в последующих исследованиях [Mihail, Asandei, 1961], показавших, что кормление мясом моллюсков активизирует гипофиз и ведет к увеличению нарастания веса головастиков на 60%. Можно полагать, что и «эффект Денеля» имеет в своей основе эндокринологический механизм. С исследованиями Денеля перекликаются выводы некоторых проведенных на амфибиях экспериментов, которые указывают на сезонную изменчивость особенностей скелета [Cohen, 1962]. У беспозвоночных животных условия среды могут явиться непосредственной причиной кардинальных изменений в морфологии и физиологии. Г. X. Шапошников [1965, 1966] показал, что при воспитании тлей Dysophis anterisci на разных кормовых растениях можно получить формы, приближающиеся к различным видам. Более того, автор утверждает, что возникшие таким путем формы теряют способность к скрещиванию, но оказываются плодовитыми при скрещивании с другими видами. Возможно, что на конечные выводы автора в какой-то мере повлияла недостаточная разработанность систематики тлей, но его исследования свидетельствуют очень ярко о громадном влиянии внешних условий на морфофизиологические особенности насекомых.
Во многих случаях внешняя среда действует как переключатель, определяющий ход развития по одному из нескольких возможных альтернативных путей. Изменение концентрации солей в среде вызывает глубокую перестройку внутриклеточной организации у Naegleria gruberi (Bistadiidac). Эти простейшие, развиваясь на сухом субстрате в присутствии бактерий, имеют амебоидную форму; развиваясь в воде, они образуют жгутики, а вся клетка удлиняется и приобретает структуру, характерную для жгутиковых [Willmer, 1956]. У пресноводной улитки Theodoxus fluviatilis встречаются три резко различных типа окраски. Оказалось [Neumann, 1959], что простое изменение внешних условий (температуры, рН и содержания солей в воде) вызывает проявление окраски того или иного типа. У парамеций незначительное изменение окружающих условий приводит к проявлению различных антигенов [Beale, 1954].
Анализируя значение подобных наблюдений, Уоддингтон [1964] пишет: «...незначительное изменение окружающих условий приводит к появлению тех или иных наследственных потенций особи. Однако у Theodoxus fluviatilis эти потенции не абсолютно исключают одна другую и могут выявляться одновременно, так что пятнистая и поперечная модели накладываются друг на друга. Кроме того, фенотипические признаки в данном случае гораздо сложнее, чем антигены Paramecium. В образовании пятнистой или поперечной окраски участвует очень большое число взаимодействующих клеточных процессов, что неизбежно связано с деятельностью многих генов. Поэтому те факторы (будь то генетические
{91}
факторы или факторы внешней среды), которые приводят к образованию той или иной окраски, по-видимому, действуют как переключатели, направляющие развитие в сторону образования одной из двух альтернативных систем, каждая из которых значительно более сложна, чем система образования одного антигена у Paramecium. Быть может, наиболее близким примером служит переключение между жгутиковой и амебоидной формами у Naegleria».
Сходные, но, пожалуй, еще более интересные наблюдения были сделаны на бабочках рода Papilio (см. обзор [Sheppard, 1959]). У Р. machaon было обнаружено, что при окукливании гусениц на листьях куколки приобретают зеленую окраску, при окукливании на коре — коричневую. Сходное происходит и у Р. polyxenae. Анализ этих наблюдений показал, что зеленые и коричневые куколки отличаются не только окраской, но и скоростью развития: большинство коричневых куколок перезимовывает, зеленые дают бабочек осенью. В данном случае мы сталкиваемся с очень интересной формой полиморфизма, который основан не на существовании в популяции двух резко различных генотипов, а на разном проявлении одного генотипа. Этот тип полиморфизма может быть назван фенотипическим полиморфизмом (environmental polymorphism [Sheppard, 1959]). Биологическое значение фенотипического полиморфизма понятно: зимовка куколок на листьях практически невозможна, и поэтому криптические (под цвет листа) куколки должны закончить превращение как можно раньше. Эти наблюдения очень ясно показывают, что простейшее изменение внешних условий (цвет субстрата) может привести к кардинальным изменениям физиологических особенностей животных (скорость развития). Характер влияния среды на организм определяется, естественно, не только природой действующего фактора, но и генетической спецификой организма (утверждение о том, что наследуются не признаки, а норма реакции, воспринимается сейчас как аксиома). Значит, одно и то же наследственное изменение вызывает различное фенотипическое действие в зависимости от того, в какой среде развивается животное. Особенно резко это проявляется в тех случаях, когда на первый план выступает так называемый вторичный эффект действия генов, суть которого заключается в том, что относительно второстепенные генетические различия влекут за собой многообразные и резко выраженные морфофизиологические следствия, которые не имеют собственной генетической основы. В качестве примера можно привести мутацию frizzled у кур. Непосредственное проявление этой мутации — скручивание перьев, вторичный эффект (связанный с нарушением нормального поддержания теплового баланса) — интенсификация метаболизма, гипертрофия желудочков аорты, увеличение общего объема крови, учащение сердцебиения, увеличение размеров кишечника, надпочечников и щитовидной железы [Landauer, 1946].
{92}
Особенности frizzled нельзя назвать фенотипическими, так как они связаны с определенной мутацией, но их трудно назвать и генетическими особенностями, так как в разной среде эффект мутации приводит к резко различным морфологическим последствиям. Вновь открытые проявления закономерностей морфофизиологических корреляций ставят перед зоологами новый круг вопросов, объединяемых общей задачей: не ограничиваться описанием отличий между сравниваемыми формами, а выяснять их приспособительную и генетическую природу. Вряд ли нужно доказывать, что решение этой задачи позволило бы мобилизовать накопленный за многие десятилетия материал, характеризующий морфологическую специфику близких форм различного таксономического ранга для решения современных проблем эволюционного учения.
Сопоставление приведенных здесь данных показывает, во-первых, что прямое влияние среды может вызвать у популяций изменения таксономического масштаба; во-вторых, межпопуляционные отличия очень редко (вероятно, никогда) не определяются или фенотипическими, или генетическими механизмами, они определяются и различиями в генетическом составе популяций, и различиями в условиях среды. Отсюда следует, что только комплексные, эколого-генетические исследования могут содействовать принципиальному прогрессу теории микроэволюции. При этом нельзя забывать, что возможны и крайние сложные случаи, анализ которых приводит к заключению о необходимости учитывать наиболее сложные механизмы эпигенетики при рассмотрении, казалось бы, очень простых природных ситуаций. С какими трудностями можно при этом столкнуться, хорошо показывает интереснейшее исследование Бернетта [Barnett, 1956], которое заслуживает того, чтобы его рассмотреть более детально.
На мясных складах мыши образуют стойкие популяции при температуре —10°. За счет физической терморегуляции мыши к столь низкой температуре приспособиться не могут (мелкие размеры тела!), и пути их адаптации к условиям жизни в холодильниках остаются невыясненными. В опытах автора лабораторные мыши воспитывались при температуре —3° в течение многих поколений. Было установлено, что в полном согласии с теорией незначительное улучшение теплоизоляционных свойств покровов не могло быть причиной их быстрой приспособляемости. Этот вывод был блестяще подтвержден и тем, что способность адаптироваться к холоду была констатирована и у мутантов, лишенных волосяного покрова. Калориметрические расчеты показали, что подопытные животные приобрели способность лучше использовать питательные вещества. Это нашло себе косвенное подтверждение в том, что лактирующие самки подопытных мышей на каждые 10 г молодняка потребляли меньше корма, чем в контроле. При разведении на холоде генетически разнородной популяции лабораторных мышей этот результат можно было бы приписать
{93}
отбору холодостойких генотипов (смертность молодняка первых двух поколений колебалась от 40 до 80%, затем она снижалась до 10% в 12-м поколении). Однако совершенно такой же результат был получен автором при работе с инбредированной линией мышей: их холодостойкость увеличилась, смертность молодняка уже в 7–10-м поколениях снизилась до 5–10%, вес тела увеличился, увеличилось содержание жира в организме. Автор считает аксиомой, что отбор в сильно инбредированной линии бессилен, и объясняет полученные результаты физиологическими особенностями матерей, приспособившихся к низким температурам (это подтвердили опыты с подменой пометов). В целом автор считает результаты опытов загадочными. В другом исследовании того же автора [Barnett, Coleman, 1960] показано, что условия среды определяют и результаты внутривидовой гибридизации: было установлено, что при скрещивании двух инбредных линий мышей гетерозис проявляется значительно сильнее, если до скрещивания животные содержались при низкой температуре.
По-видимому, Бернетт прав, результаты его опытов в самом деле загадочны, так как трудно объясняются ортодоксальными генетическими представлениями. Для их объяснения нужно, вероятно, прибегнуть к анализу данных, характеризующих феномен специфических ингибиторов [Lenick, 1963], который объясняет влияние физиологических особенностей матери, приобретенных в процессе развития в своеобразной среде, на морфофизиологические особенности потомства. Необходимо, видимо, также вспомнить и о недавно обнаруженном явлении «необычной наследственности» [Tamsitt, 1961; Dawson, 1965; Falconer, 1965], показывающем, что в определенных условиях в потомстве доминируют признаки матери (так называемый материнский эффект). Дело генетиков — разобраться в тех выводах, которые следуют из работ, подобных исследованию Бернетта. Экологу же крайне важно учитывать, что один и тот же внешний эффект может быть вызван различными генетическими механизмами, в том числе и очень сложными, комплексными. Это свидетельствует о том, что межпопуляционные различия не только безгранично разнообразны, но и основаны на практически безгранично разнообразных механизмах. Даже незначительное преобразование популяции — сложнейшее биологическое явление. Представление о том, что в одних случаях оно основано на генетико-автоматических механизмах, в других — на действии индивидуального естественного отбора, в третьих — на действии отбора группового, в четвертых — на фенотипических механизмах, в пятых — на ассимиляции приобретенных признаков (принцип Уоддингтона) и т. п., не выдерживает критики. Логично полагать, что в каждом акте преобразования популяции участвуют различные механизмы в разном сочетании. Их совместное действие и приводит к тому эффекту, который мы непосредственно фиксируем,— к преобразованию популяции.
{94}
Поэтому представляется важным детально проанализировать механизмы, приводящие к изменению в процессе онтогенеза признаков, которые до последнего времени считались наиболее стабильными, не изменяющимися при изменении условий среды.
Многие наиболее существенные особенности таксонов разных рангов (от подвидов до семейств и отрядов) определяются закономерностями роста животных, определяющими формирование их важнейших особенностей. Наиболее ярким проявлением этих закономерностей является аллометрический рост, определяющий морфологические различия между животными разных размеров. Проблеме аллометрического роста посвящены сотни работ. Однако исследования описательного направления резко преобладают над экспериментальными. Это объясняет парадоксальную по существу ситуацию: несмотря на громадный накопленный в литературе эмпирический материал, ни в одной из работ (в том числе и теоретических обобщениях) нет даже попытки объяснить те физиологические и морфогенетические механизмы, которые определяют аллометрический эффект и формируют фенотип животного. С другой стороны, изучение аллометрического роста уже со времен Т. Хаксли быстро оформилось в самостоятельное направление. Это несомненно способствовало углубленному изучению проблемы, но вместе с тем привело к известной ее изоляции, к определенному отрыву от других направлений, изучающих рост с других (отнюдь не альтернативных) теоретических и экспериментальных позиций.
Интересуясь экологической детерминированностью явлений морфогенеза, в нашей лаборатории около 20 лет тому назад были поставлены исследования по изучению общих закономерностей роста позвоночных животных. Исследования развивались в следующих направлениях: экспериментальное изучение явлений аллометрии (основные работы проведены В. Г. Ищенко); изучение взаимосвязи морфогенеза животных со скоростью их роста; изучение биологических особенностей сезонных генераций животных, характеризующихся разной скоростью роста (основные работы проведены нами, А. В. Покровским, В. Г. Оленевым); экспериментальное изучение скорости роста и развития личинок земноводных (основные исследования О. А. Пястоловой). Результаты работ в указанном направлении нашли отражение в многочисленных публикациях, здесь же мы даем их синтез.
Основные выводы работ по экспериментальному изучению аллометрии могут быть проиллюстрированы исследованием, проведенным при сопоставлении двух подвидов узкочерепной полевки [Ищенко, 1966]. Для сравнения был использован материал из вивария нашей лаборатории, так как различия, проявляющиеся между подвидами в равных условиях, с большим основанием можно рассматривать как наследственные, а не модификационные. Просмотрено было 144 черепа М. gregalis gregalis, 228 черепов М. g. major и 68 черепов их гибридов (F1). Кроме того, автор
{95}
воспользовался промерами 587 черепов М. g. major от животных, добытых К. И. Копейным на п-ове Ямал в 1956–1957 гг.
Сравнение роста отдельных частей черепа (длины зубного ряда, скуловой ширины, ширины межглазничного промежутка, высоты черепа) по отношению к кондилобазальной длине черепа, а также относительного роста размеров черепа и длины тела показало, что изучаемые подвиды в условиях вивария характеризуются специфическими аллометрическими зависимостями (см. табл. 7). Это еще раз подтверждает, что морфологические различия между данными подвидами наследственны, а не обусловлены действием внешних факторов. Гибриды этих подвидов по абсолютным значениям признаков занимают промежуточное положение [Шварц и др., 1960]. Однако, как видно из табл. 7, это промежуточное положение достигается различными путями. Для величины межглазничного промежутка свойственно промежуточное значение а и b, следовательно, данный признак, как, впрочем, и все наследуемые промежуточно в F1, является полигенным, а полигенность, как известно,— один из надежных способов защиты нормального формообразования [Шмальгаузен, 1964].
<![if !vml]>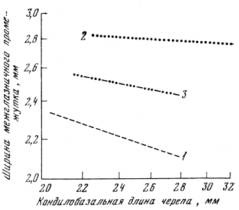 <![endif]>
<![endif]>
Рис. 3. Изменение ширины межглазничного промежутка (в мм) относительно размеров черепа у Microtus gregalis gregalis (1), М. g. major (2) и их гибридов (3)
В основном М. g. gregalis и М. g. major характеризуются близкими значениями а, но различными значениями параметра b. Это наблюдается при сравнении относительного роста кондилобазальной длины черепа, высоты и скуловой ширины его, т. е. тех частей, для которых характерна обычная аллометрия (положительная или отрицательная). Однако что касается ширины межглазничного промежутка, то изучение его роста показало, что у М. g. major величина этой части при изменении размеров черепа почти не меняется (а= -0,041), а у М. g. gregalis для этого признака характерна энантиометрия (а= -0,319), т. е. с увеличением размеров черепа величина межглазничного промежутка убывает (рис. 3). В абсолютном выражении межглазничный промежуток у М. g. major значительно шире, чем у М. g. gregalis.
{96}
Возникает вопрос: каким образом при дифференцировке вида Microtus gregalis и при образовании М. g. major изменилась аллометрическая зависимость — понадобилось ли при этом дополнительное мутирование, изменившее скорость роста, или для отбора было достаточно вариантов, имеющихся в популяции М. g. gregalis? Анализ нашего материала показал, что особи М. g. gregalis, обладающие наибольшим значением межглазничного промежутка для данного подвида (2,5–2,6 мм) и составляющие в лабораторной популяции 10,5%, характеризуются почти тем же аллометри-ческим показателем межглазничного промежутка, что и М. g. major (а — 0,080). Из этого можно предположить, что в процессе выделения подвида М. g. major для «слома» аллометрической зависимости достаточно было имеющегося в пределах М. g. gregalis многообразия, тем более что по этому признаку между данными подвидами хиатуса нет.
Но главное внимание мы считаем необходимым обратить на следующее. Если бы при расселении М. g. gregalis на север отбор шел только в направлении увеличения размеров тела и черепа, без изменения аллометрической зависимости между межглазничным промежутком и кондилобазальной длиной черепа, то в силу обратной корреляции между этими признаками у М. g. gregalis абсолютное значение межглазничного промежутка у М. g. major оказалось бы настолько мало, что его явно не хватило бы для нормального функционирования черепа. Экстраполируя алломет-рическое уравнение, мы получили, что при простом увеличении черепа М. g. gregalis у крупных особей М. g. major ширина межглазничного промежутка должна бы составлять 1,8 мм и менее, т. е. череп был бы весьма подвержен механическим повреждениям. Следовательно, отбор по одному признаку (размеры черепа) невозможен и мог идти лишь в направлении одновременного изменения ряда признаков, в том числе и величины межглазничного промежутка, причем основным материалом для него, на наш взгляд, является внутрипопуляционная изменчивость М. g. gregalis.
Интересно, что в дикой популяции М. g. major обнаруживается значительная индивидуальная изменчивость ширины межглазничного промежутка (от 2,4 до 3,4 мм, особи со значением межглазничного промежутка более 3 мм составляют 21,6% ямальской популяции). Сравнивая аллометрические зависимости животных, добытых осенью 1956 г. и ранней весной 1957 г., мы получили различные аллометрические зависимости для этого признака. Для животных, добытых осенью 1956 г., зависимость между шириной межглазничного промежутка и кондилобазальной длиной черепа выражается уравнением
у=11,84х–0,430 (n=121),
а для животных, добытых весной 1957 г.,— уравнением
у=4,50х–0,144 (n=124).
{97}
Таблица 7
Значения параметров аллометрических уравнений для двух подвидов узкочерепной полевки и их гибридов
|
Признак |
М. g. gregalis |
М. g. major |
М. g. gregalis X М. g. major, F1 |
|||
|
|
b |
a |
b |
a |
b |
a |
|
Кондилобазальная длина черепа Ширина межглазничного промежутка Скуловая ширина Длина зубного ряда Высота черепа |
3,990 6,140 0,249 1,014 1,980 |
0,394 -0,319 1,200 0,522 0,436 |
3,500 3,240 0,244 0,587 1,790 |
0,431 -0,041 1,216 0,698 0,476 |
2,500 4,470 0,167 1,053 2,076 |
0,501 -0,178 1,320 0,520 0,430 |
Однако в состав животных, добытых весной, входят особи, родившиеся под снегом, развивающиеся в иных условиях, чем добытые осенью. Это и могло быть причиной изменения аллометрической зависимости. Однако если взять весной только перезимовавших особей (n=32), то эта зависимость будет иметь вид y=6,78x–0,266, т. е. тоже отличный от зависимости, характеризующей животных, добытых осенью. Особи, добытые осенью и перезимовавшие, развивались в равных условиях, и поэтому различия в аллометрических зависимостях в данном случае могут быть объяснены элиминацией животных в течение зимнего периода, сменой генотипического состава популяции. Аналогично можно предположить, что с продвижением на север М. gregalis крупные особи с наибольшим межглазничным промежутком сохранялись, обратные же варианты элиминировались, что в конце концов и привело к изменению аллометрической зависимости.
Если объединить особей М. g. major, добытых в природе в разные годы (а при обработке виварного материала использовались животные, забитые в разное время в течение трех лет), то зависимость между межглазничным промежутком и кондилобазальной длиной черепа выразится уравнением у=10,18х–0,391, т. е. величина а будет сходна с таковой у М. g. gregalis, а параметр b отличен (Р<0,001). Это еще раз подтверждает, что отбор при выделении подвида М. g. major мог идти только в направлении одновременного изменения как размеров, так и пропорций за счет имеющегося в пределах М. g. gregalis многообразия. По-видимому, для форм, характеризующихся наличием «отрицательного роста» каких-либо частей, отбор в направлении изменения только размеров тела невозможен.
Изменение аллометрических зависимостей свидетельствует о далеко зашедшей морфологической дифференциации, приведшей к генетической разнокачественности М. g. gregalis и М. g. major.
{98}
Гибридизация этих подвидов также подтверждает генетическую разнокачественность их. Изучение аллометрического роста черепа по отношению к размерам тела показало, что для гибридов характерен иной тип аллометрической зависимости, не промежуточный и не имеющийся у родительских форм (рис. 4): мелкие гибридные особи имеют череп такой же величины, как мелкие М. g. gregalis а крупные гибриды характеризуются индексом черепа, свойственным крупным М. g. major. Это представляет большой интерес и по-видимому, связано с изменением скорости роста у гибридных особей. Во всяком случае, генетическая разнокачественность родительских форм обусловила то, что при смешении разных генотипов возникли новые, а это нашло фенотипическое отражение в изменении аллометрической зависимости [Ищенко, 1966].
<![if !vml]>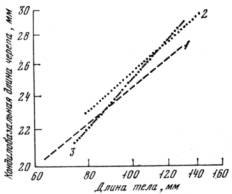 <![endif]>
<![endif]>
Рис. 4. Рост черепа относительно общих размеров у Microtus gregalis gregalis (1), М. g major (2) и их гибридов (3)
В заключение подчеркнем: можно считать экспериментально доказанным, что фактор b при изменении условий существования животных резко изменяется, а остается в пределах генетической «нормы» даже при резких изменениях жизни животных, но у животных генетически своеобразных изменяется. Экологическая целесообразность этих изменений доказана. Оставалось, однако, не ясным, могут ли быть преодолены законы аллометрии при изменении фундаментальных физиологических особенностей животных. Наблюдения в природе давали основания полагать, что ответ на этот вопрос может быть получен при изучении морфологических особенностей животных, характеризующихся разной скоростью роста [Шварц, 1961б].
Исследования, проведенные на многих видах полевок, показали, что сезонные генерации характеризуются комплексом биологических особенностей. Грызуны, родившиеся весной, быстро растут, достигают половой зрелости в возрасте около месяца (у полевки-экономки и узкочерепной полевки известны случаи полового созревания в возрасте 9–10 дней), приносят несколько пометов молодняка и к осени (в возрасте 5–6 месяцев) вымирают со всеми признаками старости. Этому заключению не противоре-
{99}
чат данные, свидетельствующие о том, что в лабораторных условиях виды полевок могут жить до трех лет, сохраняя в состоянии явной старческой дряхлости способность к размножению.
Осенние генерации грызунов после краткого периода ювенильного роста прекращают расти; скорость клеточного деления существенно снижается, у корнезубных полевок прекращается рост коренных зубов; скорость нарастания веса хрусталика глаза (один из наиболее универсальных признаков старения) резко снижается, инволюция тимуса прекращается. Весной грызуны, родившиеся во второй половине лета предшествующего года, в возрасте 9–11 месяцев сохраняют все признаки физиологической юности. Возобновление роста весной совпадает с увеличением веса тимуса и с началом полового созревания. Их дальнейшее развитие практически совпадает с развитием грызунов весенней генерации, которые младше их почти на год.
Эти данные однозначно свидетельствуют о том, что скорость процесса старения млекопитающих не фиксирована наследственностью в жестких пределах и может быть увеличена в 2–3 раза, и, что наиболее важно, не за счет продления заключительных этапов онтогенеза (активная старость), а за счет увеличения продолжительности юности. Общий вывод проведенных исследований в нашей лаборатории обосновывается значительным материалом. Он был получен при изучении многих видов полевок и мышей, в том числе и на представителях субарктической фауны. В последнем случае у полевок фаза «физиологической юности» растягивается почти на целый год [Пястолова, 1971]. Общие выводы этих исследований были подтверждены на американских грызунах и насекомоядных. Ряд физиологических особенностей осенних генераций, естественно, связывается с торможением процессов старения. Среди них по понятным причинам особое значение имеют снижения уровня обмена и замедленный темп клеточного деления. Однако полностью приписать различия между генерациями указанным особенностям рискованно.
В связи с рассматриваемым вопросом особый интерес представляют данные, показывающие, что разные генерации отличаются различной скоростью роста и различными пропорциями тела. С наибольшей полнотой этот вопрос был изучен на узкочерепной полевке [Шварц и др., 1960]. Сравнивались два подвида — М. g. gregalis Pall. и М. g. major Ogn. Было установлено, что наряду с окраской различия в общих размерах тела — одно из основных отличий между сравниваемыми формами. Насколько существенны эти различия, видно из табл. 8, в которой представлены данные, характеризующие максимальные размеры тела и черепа полевок обоих подвидов.
Сложнее устанавливаются различия в пропорциях тела и черепа. Дело в том, что, как показали исследования, проведенные в нашей лаборатории, пропорции тела и черепа грызунов зависят де только от абсолютных размеров животных (аллометрический
{100}
Таблица 8
Максимальные размеры тела и черепа (в мм) М. g. gregalis и М. g. major
|
Подвид |
Вес тела |
Длина тела |
Кондилобазальная длина |
|
М. g. gregalis (Курганская область) |
49,7 |
120,0 |
27,1 |
|
М. g. gregalis (виварий) |
42,5 |
120,0 |
26,9 |
|
М. g. major (п-ов Ямал) |
67,7 |
148,0 |
30,0 |
|
М. g. major (виварий) |
86,5 |
137,0 |
30,0 |
рост), но и от условий, в которых проходят их рост и развитие, от скорости их роста.
Так как условия развития и скорость роста животных в разное время года различны, можно было бы ожидать, что пропорции тела животных, добытых в разное время года, будут различными даже и в том случае, когда для сравнения подбираются особи одинаковых размеров.
Таблица 9 показывает, что эти различия проявляются отчетливо при анализе природных популяций. Они настолько значительны, что заставляют сомневаться в реальности различий между подвидами не только в тех случаях, когда сравниваемые формы представлены особями, отличающимися по размерам, но и когда они добыты в разное время года. Из таблицы видно, что в течение года происходит вполне закономерное изменение в пропорциях тела и черепа животных, которое наблюдается во всех размерных и возрастных группах. При этом очень важно отметить, что различия в пропорциях тела и черепа животных одного размера, но добытых в разное время года, могут быть больше, чем у животных разных размеров, но добытых одновременно. Отношение скуловой ширины к кондилобазальной длине черепа с увеличением размеров тела животных закономерно увеличивается. Однако у полевок весом от 10 до 20 г, добытых в июле, относительная скуловая ширина черепа больше, чем у полевок весом 30–40 г, добытых весной.
В табл. 10 приведены сравнительные данные, характеризующие пропорции тела и черепа старых особей двух подвидов полевок. Из них видно, что у М. g. major хвост короче, задняя ступня длиннее, относительная длина лицевой части черепа больше, межглазничный промежуток значительно шире, высота черепа меньше. Большинство этих различий проявляется и при сравнении животных других размеров (табл. 11).
Однако эти различия могут быть выявлены лишь при соблюдении тех условий сравнения, о которых мы говорили выше. Следует поэтому особо подчеркнуть, что если сравнивать животных
{101}
Таблица 9
Изменение пропорций тела и черепа M.g. major в зависимости от общих размеров и времени добычи животных
|
Показатель |
Весовая группа, г |
|||||||||||
|
10—20 |
20—30 |
30—40 |
||||||||||
|
Число обследованных особей |
||||||||||||
|
3 |
6 |
12 |
27 |
15 |
13 |
35 |
29 |
20 |
22 |
11 |
49 |
|
|
Время добычи |
||||||||||||
|
15.IV–20.V |
1—20.VI |
1—15.VII |
1–30.VIII |
10.IX—15.Х |
15.IV—20.V |
1—20.VI |
1—15.VII |
1—30.VIII |
10.IX—15.Х |
15.IV—20.V |
20.VI |
|
|
Отношение скуловой ширины к кондило-базальной длине черепа |
0,474 |
0,499 |
0,520 |
0,497 |
0,495 |
0,496 |
0,498 |
0,492 |
0,497 |
0,503 |
0,508 |
0,519 |
|
Отношение длины лицевой части черепа и мозговой |
1,49 |
1,61 |
1,47 |
1,58 |
1,53 |
1,51 |
1,58 |
1,44 |
1,49 |
1,50 |
1,15 |
1,53 |
|
Отношение ширины межглазничного промежутка к кондило-базальной длине черепа |
0,108 |
0,112 |
0,144 |
0,134 |
0,133 |
0,107 |
0,111 |
0,135 |
0,129 |
0,125 |
0,104 |
0,105 |
|
Отношение наибольшей высоты черепа к кон-дилобазальной длине |
0,331 |
0,342 |
0,411 |
0,384 |
0,367 |
0,336 |
0,342 |
0,347 |
0,372 |
0,362 |
0,325 |
0,335 |
|
Отношение кондилоба-зальной длины к длине тела |
0,266 |
0,242 |
0,239 |
0,263 |
0,271 |
0,253 |
0,234 |
0,215 |
0,246 |
0,258 |
0,236 |
0,226 |
|
Отношение длины хвоста к длине тела |
0,222 |
0,212 |
0,194 |
0,241 |
0,233 |
0,228 |
0,215 |
0,208 |
0,223 |
0,223 |
0,203 |
0,207 |
|
Отношение длины ступни к длине тела |
0,178 |
0,163 |
0,182 |
0,191 |
0,168 |
0,157 |
0,157 |
0,165 |
0,175 |
0,182 |
0,155 |
0,152 |
{102}
Таблица 9 (окончание)
<![if !supportMisalignedRows]><![endif]><![if !supportMisalignedRows]><![endif]><![if !supportMisalignedRows]><![endif]><![if !supportMisalignedRows]><![endif]><![if !supportMisalignedRows]><![endif]><![if !supportMisalignedRows]><![endif]><![if !supportMisalignedRows]><![endif]><![if !supportMisalignedRows]><![endif]><![if !supportMisalignedRows]><![endif]><![if !supportMisalignedRows]><![endif]><![if !supportMisalignedRows]><![endif]><![if !supportMisalignedRows]><![endif]><![if !supportMisalignedRows]><![endif]><![if !supportMisalignedRows]><![endif]>
|
Показатель |
Весовая группа |
|
|||||||||||
|
30—40 |
40—50 |
50 и выше |
|
||||||||||
|
Число обследованных особей |
|
||||||||||||
|
35 |
17 |
17 |
19 |
17 |
15 |
3 |
2 |
7 |
24 |
5 |
1 |
|
|
|
Время добычи |
|
||||||||||||
|
1—15.VII |
1—30.VIII |
10.IХ—15.Х |
1—20.VI |
1—15.VII |
1—30.VIII |
10.IX—15.Х |
15.IV—20.VI |
1—20.VI |
1—15.VII |
1—30.VIII |
10.IX—15.Х |
||
|
Отношение скуловой ширины к кондилоба-зальной длине черепа |
0,529 |
0,528 |
0,525 |
0,525 |
0,527 |
0,534 |
0,534 |
0,526 |
0,537 |
0,539 |
0,534 |
0,564 |
|
|
Отношение длины лицевой части черепа к мозговой |
1,46 |
1,51 |
1,54 |
1,55 |
1,49 |
1,55 |
1,52 |
1,59 |
1,52 |
1,50 |
1,52 |
1,54 |
|
|
Отношение ширины межглазничного проме-жутка к кондило-базальной длине черепа |
0,123 |
0,122 |
0,107 |
0,104 |
0,109 |
0,105 |
0,099 |
0,097 |
0,098 |
0,101 |
0,093 |
0,098 |
|
|
Отношение наибольшей высоты черепа к кондилобазальной длине |
0,365 |
0,347 |
0,344 |
0,335 |
0,351 |
0,341 |
0,329 |
0,325 |
0,326 |
0,333 |
0,319 |
0,331 |
|
|
Отношение кондилоба-зальной длины к длине тела |
0,207 |
0,227 |
0,233 |
0,218 |
0,192 |
0,224 |
0,234 |
0,228 |
0,213 |
0,204 |
0,214 |
0,231 |
|
|
Отношение длины хвоста к длине тела |
0,207 |
0,251 |
0,244 |
0,194 |
0,209 |
0,230 |
0,232 |
0,224 |
0,208 |
0,206 |
0,234 |
0,153 |
|
|
Отношение длины ступни к длине тела |
0,151 |
0,152 |
0,156 |
0,146 |
0,138 |
0,147 |
0,150 |
0,152 |
0,141 |
0,132 |
0,136 |
0,153 |
|
{103}
Таблица 10
Основные показатели пропорций тела и черепа М. g. gregalis и М. g. major
(старшая возрастная группа, вес тела 40-50 г, животные добыты в августе)
|
Показатель |
М. g. major |
М. g. gregalis |
|
Отношение скуловой ширины к кондилобазальной длине черепа Отношение лицевой части черепа к мозговой Отношение ширины межглазничного промежутка к кондилобазальной длине черепа Отношение кондилобазальной длины к длине тела Отношение длины хвоста к длине тела Отношение длины ступни к длине тела |
0,534 1,55 0,105 0,224 0,230 0,147 |
0,498 1,40 0,088 0,223 0,250 0,142 |
Примечание. Было исследовано 15 экземпляров М. g. major и 8—М. g. gregalis.
без учета их размеров и времени добычи, то даже по тем признакам, по которым М. g. major и М. g. gregalis отличаются особенно отчетливо, между ними могут не обнаружиться различия. Это может быть проиллюстрировано следующим примером. Отличия в длине ступни у рассматриваемых подвидов очень велики, но даже при сравнении серии одинаковых по размерам животных между ними не обнаружится различий, если М. g. major будут собраны в июне, а М. g. gregalis — в сентябре. Если М. g. gregalis собраны весной, а М. g. major — осенью, то между ними трудно будет обнаружить отличия и в ширине межглазничного промежутка.
Нам представляется, что приведенные здесь наблюдения дают основания для утверждения, что, несмотря на резкие морфологические различия между двумя подвидами, в определенных условиях развития у каждого из них могут развиваться признаки, идентичные признакам другого подвида. Существо различий между двумя резко дифференцированными формами заключается в закономерностях их морфогенеза.
Наши суждения об особенностях помесных животных по рассматриваемым в этом разделе признакам основываются на обследовании 17 зверьков. Соответствующий материал представлен в табл. 12. Его сопоставление с материалами, характеризующими важнейшие признаки родительских форм, показывает, что по тем признакам, по которым М. g. gregalis и М. g. major отличаются вполне отчетливо, помеси занимают промежуточное положение. Это свидетельствует о генетической (полигенной) закрепленности особенностей морфогенеза животных.
Существенные отличия М. g. gregalis от М. g. major проявляются в размерах. В их основе лежат особенности роста сравнивае-
{104}
мых форм. Изучение скорости роста значительного числа животных в условиях вивария показывает, что эти различия между ними статистически реальны и весьма значительны (исследования А. В. Покровского). Из сводной диаграммы (рис. 5) видно, что в равных условиях содержания М. g. major к месячному возрасту обгоняют в весе животных южного подвида в среднем на 6 г, т. е. примерно в 1,5 раза.
Как известно, скорость роста грызунов находится в очень тесной зависимости от времени их рождения. Эта зависимость проявляется и при содержании животных в виварии. Для нас представляют особый интерес те различия, которые проявляются в этом отношении между разными формами. При сопоставлении рис. 6 и 7 видно, что разница в скорости роста между М. g. gregalis и М. g. major осенью (в сентябре) значительно более существенна, чем летом (в июне). Это является следствием различий в сезонной изменчивости скорости роста сравниваемых форм. В июне скорость роста М. g. gregalis значительно выше, чем в сентябре. Различия же июньских и сентябрьских М. g. major не превышают, тех различий, которые могут быть обнаружены между двумя наудачу взятыми группами особей одного времени рождения. Это значит, что М. g. major характеризуется не только большей скоростью роста, но и относительно большим его постоянством, относительно меньшей зависимостью от сезонной смены условий среды.
Помеси по скорости роста занимают обычно промежуточное положение между родительскими формами. Ни одна из 21 помес-
Таблица 11
Пропорции тела и черепа М. gregalis сравниваемых подвидов
(младшая возрастная группа, вес тела 10–20 *)
|
Время добычи |
Отношения |
|||||||||
|
скуловой ширины к кондилобазальной длине черепа |
длины лицевой части черепа к мозговой |
ширины межглазничного промежутка к кондило-базальной длине черепа |
|
|||||||
|
M.g. major |
M. g. gregalis (Курганская обл.) |
M.g. major |
M. g. gregalis (Курганская обл.) |
M.g. major |
M. g. gregalis (Курганская обл.) |
|
||||
|
Байдарацкая губа |
Обская губа |
Байдарацкая губа |
Обская губа |
Байдарацкая губа |
Обская губа |
|
||||
|
Июнь–июль |
0,509 |
0,546 |
0,491 |
1:54 |
1,50 |
1,48 |
0,128 |
0,147 |
0,115 |
|
|
Август |
0,497 |
0,496 |
0,455 |
1,58 |
1,54 |
1,41 |
0,134 |
0,125 |
0,108 |
|
|
Сентябрь–октябрь |
0,495 |
0,485 |
0,446 |
1,53 |
1,52 |
1,44 |
0,133 |
0,121 |
0,096 |
|
|
|
|
|
|
|
|
|
|
|
|
|
* Для таблицы использованы 65 особей М. g. major с побережья Байдарацкой губы (в июне–июле. — 18, в августе — 27, в сентябре–октябре — 15), 61 — с побережья Обской губы (соответственно 7, 10 и 44), 23 особи М. g. gregalis — из Курганской обл. (3, 3 и 17).
{105}
ной полевки, рост которых в сопоставимых условиях удалось проследить в течение нескольких месяцев, не имела средней скорости роста животных северного подвида, родившихся в одном с ними месяце.
Подводя итог проведенной работе, подчеркнем, что животные, родившиеся в разное время года, характеризуются не только разной конституцией, но и разными закономерностями ее формирования. Рассмотренные материалы служат этому разносторонней аргументацией. Поэтому лишь для иллюстрации выделим из табл. 9 следующие данные. М. g. major, добытые ранней весной в рассмотренных размерных группах, характеризуются следующим отношением длины черепа к длине тела: 0,266; 0,253; 0,236; 0,228. Идеальное подтверждение классического примера: с увеличением размера тела относительные размеры головы снижаются. Но животные самой меньшей группы, добытые в середине лета, характеризуются таким же индексом черепа (0,239), что и самые крупные животные, добытые поздней осенью. Так как использованный пример служит обязательной иллюстрацией аллометрии не только в зоологической, но и в антропологической литературе, мы считаем себя вправе подвести итог в необычной для зоолога форме: изменение условии развития приводит к тому, что младенец приобретает конституцию старика, а старик — младенца. Совокупность приведенных данных дает основания полагать, что этот своеобразный парадокс развития в первую очередь определяется скоростью роста животных. Важность вопроса определила целесообразность постановки специальных исследований.
|
Отношения |
||||||||||||
|
|
наибольшей высоты черепа к кондилобазальной длине |
кондилобазальной длины черепа к длине тела |
длины хвоста к длине тела |
длины ступни к длине тела |
||||||||
|
|
M.g. major |
M. g. gregalis (Курганская обл.) |
M.g.major |
M. g. gregalis (Курганская обл.) |
M.g. major |
M. g. gregalis (Курганская обл.) |
M.g. major |
M. g. gregalis (Курганская обл.) |
||||
|
|
Байдарацкая губа |
Обская губа |
Байдарацкая губа |
Обская губа |
Байдарацкая губа |
Обская губа |
Байдарацкая губа |
Обская губа |
||||
|
|
0,377 |
0,450 |
0,398 |
0,241 |
0,237 |
0,249 |
0,203 |
0,203 |
0,251 |
0,172 |
0,180 |
0,133 |
|
|
0,384 |
0,378 |
0,349 |
0,263 |
0,253 |
0,266 |
0,241 |
0,241 |
0,284 |
0,192 |
0,180 |
0,166 |
|
|
0,367 |
0,359 |
0,340 |
0,271 |
0,279 |
0,285 |
0,233 |
0,227 |
0,265 |
0,191 |
0,188 |
0,170 |
{106}
Таблица 12
Основные показатели пропорций тела и черепа помесей М. g. major X М. g. gregalis
<![if !supportMisalignedColumns]><![endif]>
|
Показатель |
Июнь |
Июль |
|
|||||||
|
Возрастные группы (вес тела в г) |
|
|||||||||
|
10—20 |
20—30 |
50 и выше |
10—20 |
30—40 |
40—50 |
|
||||
|
Отношение скуловой ширины к кондилобазальной длине черепа |
0,497 |
0,471 |
0,518 |
0,489 |
0,480 |
0,517 |
|
|||
|
Отношение лицевой части черепа к мозговой |
1,56 |
1,48 |
1,36 |
1,46 |
1,43 |
1,52 |
|
|||
|
Отношение ширины межглазничного промежутка к кондилобазальной длине черепа |
0,134 |
0,110 |
0,089 |
0,126 |
0,094 |
0,095 |
|
|||
|
Отношение наибольшей высоты черепа к кондилобазальной длине |
0,379 |
0,337 |
0,327 |
0,381 |
0,366 |
0,330 |
|
|||
|
Отношение кондилобазальной длины черепа к длине тела |
0,249 |
0,243 |
0,219 |
0,252 |
0,216 |
0,224 |
|
|||
|
Отношение длины хвоста к длине тела |
0,224 |
0,237 |
0,242 |
0,211 |
0,212 |
0,291 |
|
|||
|
Отношение длины ступни к длине тела |
0,184 |
0,148 |
0,137 |
0,182 |
0,136 |
0,135 |
|
|||
|
Показатель |
Август |
Декабрь |
|
|||||||
|
Возрастные группы (вес тела в г) |
|
|||||||||
|
10—20 |
40—50 |
10—20 |
20—30 |
|||||||
|
Отношение скуловой ширины к кондилобазальной длине черепа |
0,422 |
0,488 |
0,465 |
0,514 |
||||||
|
Отношение лицевой части черепа к мозговой |
1,45 |
1,55 |
1,47 |
1,52 |
||||||
|
Отношение ширины межглазничного промежутка к кондилобазальной длине черепа |
0,120 |
0,093 |
0,105 |
0,099 |
||||||
|
Отношение наибольшей высоты черепа к кондилобазальной длине |
0,365 |
0,318 |
0,358 |
0,361 |
||||||
|
Отношение кондилобазальной длины черепа к длине тела |
0,258 |
0,232 |
0,279 |
0,231 |
||||||
|
Отношение длины хвоста к длине тела |
0,254 |
0,255 |
0,247 |
0,271 |
||||||
|
Отношение длины ступни к длине тела |
0,192 |
0,143 |
0,200 |
0,146 |
||||||
Скорость роста органа определяется его собственным возрастом и степенью дифференцировки. С возрастом животных различия в собственном возрасте органов сглаживаются и соотносительная скорость роста частей тела животного изменяется [Шмальгаузен,
{107}
1935]. Законно полагать, что все условия, интенсифицирующие рост, будут оказывать на различные органы в соответствии со степенью их дифференцировки различное влияние 1. Изменение пропорций тела животных становится в связи с этим неизбежным. С другой стороны, это объяснение различий между животными разных генераций находит себе прямую поддержку в некоторых новейших экспериментальных исследованиях [Widdowson, Mc Cance, 1960; Dickerson, Widdowson, I960], показавших, что особенности конституции животных находятся в отчетливой связи со скоростью их роста.-
<![if !vml]> <![endif]>
<![endif]>
Рис. 5. Скорость роста Microtus gregalis major (1 и М. g. gregalis (2)
К решению поставленной задачи мы подошли двумя путями. Сопоставлялись размеры тела и черепа животных, родившихся и выросших в неволе, т. е. тех, скорость роста которых может быть точно определена. Для сравнения должны были быть использованы животные с совершенно одинаковыми размерами тела, рост которых идет с разной скоростью. Так как период роста грызунов по сравнению с общей продолжительностью их жизни очень велик, то подбор соответствующих групп животных сопряжен с большими трудностями. Эти трудности увеличиваются и в силу того, что быстро растущие животные обычно оказываются и более крупными, а пользоваться для сравнения животными, находящимися еще в периоде юношеского роста, по вполне понятным причинам нельзя. Поэтому даже в условиях большого вивариума подбор массового материала, пригодного для сравнения в интересующем нас плане, затруднен.
В нашей работе использован следующий материал, характеризующий особенности конституции животных в зависимости от скорости их роста: степные пеструшки (Lagurus lagurus) — 275 экз., полевки Миддендорфа (Microtus middendorffi) — 49, северный подвид узкочерепиой полевки (М. gregalis major) — 70 южный подвид узкочерепной полевки (М. g. gregalis) — 70, помеси
1 «Мало дифференцированная, молодая протоплазма богаче нуклеопротеидами и способна к высоким уровням синтеза роста» [Никитин, 1961].
{108}
<![if !vml]> <![endif]>
<![endif]>
Рис. 6. Скорость роста Microtus gregalis major (1) и М. g. gregalis (2) июньских (а) и сентябрьских (б) пометов
<![if !vml]> <![endif]>
<![endif]>
Рис. 7. Скорость роста Microtus gregalis major (1), М. g. gregalis (2), помесей: самец М. g. major X самка М. g. gregalis (3), самец М. g. major X самка (самец M. g. gregalis X самка M. g. major) (4), самец М. g. gregalis Х самка (самец M. g. gregalis Х самка М. g. major) (5), самец (самец М. g. gregalis Х самка М. g. major) X самка М. g. gregalis (6) июньских (а) и августовских (б) пометов
этих двух подвидов узкочерепной полевки — 95. Использован материал, полученный в виварии нашей лаборатории под руководством А. В. Покровского, советами которого мы пользовались и при анализе данных. Принцип подбора животных для сравнения ясен из приведенного ниже описания отдельных экспериментов. Суждение о достоверности выводов основывалось на биометрической обработке полученных данных.
Другой путь решения задачи заключался в изучении пропорций тела и черепа таких видов животных из природных популяций, сравнительный возраст которых может быть точно определен. В качестве объекта исследования была избрана ондатра.
В табл. 13 представлен материал, характеризующий краниологические особенности двух размерных групп степных пеструшек: крупных (длина тела 92–105 мм) и мелких (длина тела 80–85 мм). Каждая из этих групп в свою очередь подразделена
{109}
Таблица 13
Краниологические признаки (в мм) степных пеструшек в зависимости от скорости их роста
(лабораторная колония)
|
Показатель |
Длина тела |
Кондилобазальная длина черепа |
Скуловая ширина |
Ширина межглазничного промежутка |
Длина зубного ряда |
Длина лицевой части черепа |
Высота черепа |
|
Первая размерная, группа (длина тела 80–85 мм) |
|||||||
|
Группа животных, растущих медленно |
84,0±0,523 |
21,9±0,266 |
13,9±0,160 |
2,57±0,049 |
5,37±0,086 |
7,98±0,139 |
7,1±0,105 |
|
Группа животных, растущих быстро |
82,8±0,688 |
20,5±0,192 |
12,5±0,108 |
2,67±0,037 |
5,06±0,049 |
7,15±0,102 |
7,4±0,219 |
|
Достоверность различий между группами |
1,4 |
4,0 |
4,8 |
1,9 |
3,1 |
4,8 |
1,1 |
|
Отношение индексов животных сравниваемых групп |
|
0,256 |
0,612 |
0,117 |
0,245 |
0,364 |
0,322 |
|
0,247 |
0,610 |
0,130 |
0,247 |
0,348 |
0,360 |
||
|
Вторая размерная группа (длина тела 92–105 мм) |
|||||||
|
Группа животных, растущих медленно |
95,0±0,617 |
22,5±0,244 |
13,7±0,152 |
2,60±0,052 |
5,50±0,048 |
8,20±0,098 |
7,3±0,071 |
|
Группа животных, растущих быстро |
96,2±1,17 |
22,0±0,141 |
13,4±0,124 |
2,80± 0,046 |
5,30±0,049 |
7,83±0,078 |
7,6±0,066 |
|
Достоверность различий между группами |
0,91 |
1,7 |
1,4 |
3,1 |
3,1 |
3,27 |
3,1 |
|
Отношение индексов животных сравниваемых групп |
|
0,237 |
0,610 |
0,115 |
0,246 |
0,365 |
0,324 |
|
0,229 |
0,608 |
0,127 |
0,245 |
0,351 |
0,345 |
||
{110}
на медленно и быстро растущих животных. Из таблицы видно что при совершенно одинаковых размерах тела пропорции тела и черепа медленно и быстро растущих животных оказываются существенно различными.
У медленно растущих животных кондилобазальная длина черепа, скуловая ширина, длина зубного ряда и лицевой части больше, а высота черепа и ширина межглазничного промежутка меньше, чем у растущих быстро. Относительно замедленный рост черепа быстро растущих животных сопровождается и известными изменениями его пропорций. Череп их не только абсолютно, но и относительно более высокий и широкий, с менее развитой лицевой частью.
Попытаемся оценить масштабы различий между животными медленно и быстро растущими. Для этого сопоставим особенности этих двух групп с особенностями животных различных размеров определяемыми законами аллометрического роста.
В табл. 14 представлены данные, показывающие изменения пропорции тела и черепа пеструшек соответственно изменениям их размеров. По размерам черепа быстро растущие животные первой группы (длина тела 80–85 мм) отличаются от медленно растущих больше, чем «средние» пеструшки с длиной тела 86–90 мм от пеструшек 75-80 мм. Для скуловой ширины и высоты черепа отличия между сравниваемыми группами соответствуют отличиям животных длиной тела 76–80 и 91–95 мм (!). Аналогичная закономерность отчетливо проявляется и при рассмотрении второй размерной группы (длина тела 92–105 мм). Аналогия с тем что наблюдается в природных популяциях,— полная!
Эти данные показывают, что различия в скорости роста могут полностью замаскировать проявление законов аллометрии. У крупных, но медленно растущих животных индекс черепа может быть больше, чем у более мелких, но быстрее растущих.
Чтобы закончить пример с пеструшками, необходимо затронуть еще один вопрос. Как мы уже указывали, быстро растущие животные оказываются, как правило, и более крупными. Поэтому группа медленно растущих представлена в нашем материале в среднем животными более старшими, чем группа быстро растущих. В связи с этим можно было бы полагать, что отмечаемые нами различия — возрастные. Однако если часть признаков быстрорастущих особей действительно характерна для молодых животных (большая высота черепа и меньшее развитие лицевой части) то другие (меньшая относительная длина черепа) свойственны наоборот, более старым животным.
То, что отдельные признаки животных рассматриваемых нами групп в известной степени определяются их возрастом, несомненно, но данные свидетельствуют и о самостоятельном влиянии скорости роста. Этот вывод подтверждается следующими наблюдениями. Даже из очень большого материала (в нашем виварии ежегодно проходят через различные эксперименты несколько ты-
{111}
Таблица 14
Краниологические особенности (в мм) степных пеструшек в зависимости от размеров тела
(лабораторная колония)
|
Размерная группа (длина тела, мм) |
Длина черепа |
Скуловая ширина |
Ширина межглазничного промежутка |
Длина зубного ряда |
Длина лицевой части черепа |
Высота черепа |
||||||
|
71–75 |
19,7±0,213 |
(21) |
12,1±0,128 |
(21) |
2,6±0,059 |
(21) |
4,9±0,047 |
(21) |
6,8±0,111 |
(21) |
7,0±0,066 |
(21) |
|
0,268 |
0,614 |
0,132 |
0,249 |
0,345 |
0,355 |
|||||||
|
76–80 |
20,7±0,185 |
(29) |
12,6±0,092 |
(29) |
2,6±0,033 |
(29) |
5,0±0,054 |
(29) |
7,4±0,087 |
(29) |
7,1±0,070 |
(29) |
|
0,264 |
0,608 |
0,125 |
0,241 |
0,357 |
0,343 |
|||||||
|
81–85 |
21,4±0,215 |
(29) |
13,0±0,133 |
(29) |
2,6±0,031 |
(29) |
5,2±0,058 |
(29) |
7,7±0,087 |
(29) |
7,2±0,055 |
(29) |
|
0,256 |
0,607 |
0,121 |
0,242 |
0,360 |
0,326 |
|||||||
|
86–90 |
21,5±0,134 |
(43) |
13,2±0,110 |
(41) |
2,6±0,026 |
(43) |
5,3±0,039 |
(43) |
7,7±0,071 |
(43) |
7,2±0,55 |
(41) |
|
0,243 |
0,613 |
0,120 |
0,246 |
0,357 |
0,335 |
|||||||
|
96–100 |
22,2±0,148 |
(42) |
13,5±0,107 |
(39) |
2,7±0,032 |
(41) |
5,4±0,043 |
(42) |
8,0±0,063 |
(42) |
7,4±0,041 |
(37) |
|
0,228 |
0,613 |
0,120 |
0,240 |
0,360 |
0,333 |
|||||||
|
101–105 |
22,9±0,135 |
(17) |
14,1±0,152 |
(17) |
2,7±0,057 |
(17) |
5,5±0,0б5 |
(17) |
8,3±0,096 |
(17) |
7,5±0,062 |
(17) |
|
0,221 |
0,617 |
0,112 |
0,241 |
0,375 |
0,320 |
|||||||
|
106–114 |
24,0±0,290 |
(9) |
14,8±0,183 |
(9) |
2,7±0,048 |
(9) |
5,8±0,090 |
(9) |
9,0±0,097 |
(9) |
7,7±0,108 |
(8) |
|
0,221 |
0,617 |
0,112 |
0,241 |
0,375 |
0,320 |
|||||||
Примечание. В числителе — абсолютное значение признака в мм, в знаменателе — индекс; в скобках — 0,320 число обследованных особей.
{112}
сяч особей разных видов грызунов) трудно подобрать группу животных, которая при строго одинаковых размерах тела и одинаковом возрасте обладала бы разной скоростью роста. После тщательного отбора нам удалось подобрать 26 таких особей. В группе с длиной тела 86–95 мм у медленно растущих особей в возрасте 100–123 дней индекс высоты черепа оказался в среднем 0,322±0,003, у быстро растущих — 0,336±0,003 (достоверность различий 3,55). В группе с длиной тела 91–100 мм в возрасте 96–123 дня соответственные величины равны 0,318±0,004 и 0,322±0,003 (достоверность различий 2,86). Эти данные показывают, что даже наиболее выраженный возрастной признак (высота черепа) изменяется в зависимости от скорости роста и независимо от возраста.
О самостоятельном значении скорости роста в формировании пропорций черепа свидетельствуют и данные табл. 13 и 14. Различия между быстро и медленно растущими животными первой размерной группы в индексе высоты черепа (0,322 и 0,360), несмотря на одинаковые размеры и сопоставимый возраст, больше, чем между находящимися еще в периоде юношеского роста juvenis (длина тела 65–70 мм) и самыми крупными и старыми особями с длиной тела, превышающей 100 мм.
Анализ материала привел к заключению, что скорость роста в первый период жизни накладывает неизгладимый отпечаток на конституцию взрослых и старых животных. Изучение этой закономерности могло быть проведено путем изучения частной корреляции между размерами животных в возрасте одного месяца и развитием черепа.
Метод частного коррелирования позволяет устранить влияние какого-либо фактора (переменной) на связь других двух по формуле
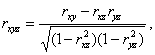
где х, у, z — соответственные переменные. Если взять три переменные величины (в нашем случае — скорость роста животного, его размеры и размеры черепа) и определить корреляции между каждой парой переменных, то появляется возможность исключить одну из них (в данном случае — размеры животного, так как их связь с размерами черепа твердо установлена) и тем самым определить корреляцию двух других переменных, которая соответствует совокупности отобранной так, что эта третья переменная оставалась бы константной.
Применив метод частного коррелирования, мы определили связь между скоростью роста в первый период жизни (выраженной в весе тела в возрасте одного месяца) и относительной длиной черепа (индекс черепа). Были получены следующие коэффициенты корреляции [r—коэффициент корреляции, р—уровень
{113}
существенности корреляции):
r p
Степная пеструшка -0,97±0,06 0,01
Узкочерепная полевка
северный подвид -0,370±0,157 0,05
южный подвид -0,08 (нет корреляции)
Полевка Миддендорфа -0,67±0,09 0,01
Как видно, у трех форм, выращенных в нашем виварии, установлена отчетливая отрицательная корреляция между весом тела в первый месяц жизни (скоростью роста) и размерами черепа. Отсутствие корреляции у южного подвида узкочерепной полевки, вероятно, связано с какими-то нарушениями в развитии, отразившимися и на закономерностях их роста 1.
При наличии большого материала, допускающего разбивку животных на группы, можно анализировать связь скорости роста животных с отдельными признаками путем вычисления соответствующих коэффициентов общей корреляции в пределах отдельных размерных групп.
Так, например, у М. gregalis major коэффициент корреляции между весом тела в первый месяц жизни и длиной лицевой части у животных первой размерной группы (длина тела 80–90 мм) равен — 0,41±0,2 (достоверность 1,84), второй группы (90–100 мм) — 0,65±0,14 (достоверность 4,6) и третьей группы (110–120 мм) — 0,59±0,27 (достоверность 2,2).
Совокупность приведенных данных показывает, что зависимость пропорций тела и черепа животных от скорости роста находит себе экспериментальное подтверждение и проявляется у всех изученных в этом отношении видов.
По понятным причинам изучение интересующей нас закономерности в природных условиях возможно только на видах, методика определения возраста которых хорошо разработана. Поэтому в качестве объекта исследования была выбрана ондатра. Для определения ее возраста мы использовали разработанную в нашей лаборатории В. С. Смирновым [Смирнов, Шварц, 1959] простую и достаточно точную методику.
Осенью 1955 г. в Лебяжьевском ондатровом хозяйстве (Курганская область) была отловлена серия молодых ондатр текущего года рождения, которых разбили на две размерные группы (по техническим причинам в качестве показателя размеров был использован вес тушки без шкурки и кишечника 2). Внутри отдель-
1 В нашем виварии в годы, когда проводились описанные эксперименты, южные узкочерепные полевки «шли» хуже других форм. Несколько раз наблюдалось прекращение их размножения и повышенная смертность.
2 Это представляло особые удобства в связи с тем, что в данном случае мы не ограничивались изучением пропорций черепа, а попытались также установить зависимость между скоростью роста животных и весом их внутренних органов.
{114}
вых размерных групп животные были разбиты на возрастные группы. Возраст ондатр оценивался по высоте коронки и длине корня зубов (чем ниже коронка и длиннее корень, тем старше животное). Достоверность этой методики была проверена нами ранее на очень большом материале и полная надежность ее была доказана.
Очевидно, что для находящихся в периоде роста животных установление их размеров и возраста создает надежные предпосылки для суждения о скорости их роста: при одинаковых размерах животные старшего возраста растут медленнее, младшего — быстрее.
Анализ данных (табл. 15) показывает, что по двум краниологическим показателям (длина диастемы и скуловая ширина) и по размерам внутренних органов (сердце, печень, почки) быстро растущие животные отчетливо отличаются от медленно растущих. При этом необходимо обратить внимание, что группа растущих медленно представлена в нашем материале несколько более крупными животными, размеры их органов меньше, чем у растущих быстро, что значительно увеличивает надежность выводов.
Таким образом, результаты полевых и лабораторных наблюдений принципиально совпадают: скорость роста определяет морфологические различия между животными одинаковых размеров и сопоставимого возраста. Это совпадение идет, однако, и дальше: особенности животных, характеризующихся определенной скоростью роста, в природе и в эксперименте совпадают. В обоих случаях быстро растущие животные отличаются меньшей скуловой шириной и более короткой лицевой частью черепа.
Заслуживает также внимания полное совпадение результатов наших исследований с полученными позднее экспериментальными данными [Widdowson, Mc Cance, 1960], показавшими, что увеличение скорости роста влечет за собой относительное увеличение размеров сердца.
Приведенные материалы позволяют заключить, что пропорции тела и черепа животных в значительной степени определяются скоростью их роста. Это приводит к совершенно закономерным морфологическим различиям между медленно и быстро растущими животными одинаковых размеров. Скорость роста формирует фенотип животного. Однако на фоне общих закономерностей отчетливо проявляется и специфика отдельных форм. Так, хотя в пределах подвида М. g. major относительная длина черепа и длина лицевой части связаны со скоростью роста отчетливой обратной корреляцией, быстро растущая форма (М. g. major) по этим признакам от медленно растущей формы (М. g. gregalis) не отличается. Это значит, что сравниваемые формы отличаются не только скоростью роста, но и закономерностями, управляющими формированием характерной конституции животных. Наше понимание существа различий между изучаемыми подвидами в данном случае углубляется.
<![if !supportEmptyParas]> <![endif]>