
Проблемы Эволюции |
М.: Наука, 1980
{172}
Таблица 17
Изменения аллометрического показателя (в мм) в популяции Arvicola terrestris
|
Признак |
Год |
Весна |
Осень |
|
Длина черепа |
1952 1953 1954 |
0,480± 0,008 0,371±0,011 0,508±0,010 |
0,585±0,014 0,460±0,007 0,486±0,016 |
|
Длина зубного ряда |
1952 1953 1954 |
0,354±0,012 0,785±0,014 0,629±0,016 |
0,371±0,013 0,б84±0,009 0,б18±0,025 |
|
Ширина межглазничного промежутка |
1952 1953 1954 |
-0,179±0,006 0,013±0,029 -0,192±0,029 |
0,115±0,022 -0,475±0,037 -0,394±0,026 |
|
Скуловая ширина |
1952 1953 1954 |
0,760±0,017 0,793±0,018 0,793±0,016 |
0,885±0,012 1,155±0,011 1,050±0,013 |
|
Высота мозговой части |
1952 1953 1954 |
0,768±0,019 0,695±0,054 0,568±0,034 |
0,515±0,017 0,640±0,009 0,571±0,011 |
Таблица 18
Аллометрический показатель межглазничной ширины черепа у различных видов кошек
[по Rohrs, 1959]
|
Вид |
a |
b |
Число черепов |
|
Felis ocreata |
1,052 |
0,176 |
14 |
|
F. lynx |
1,238 |
0,083 |
17 |
|
Panthera pardus |
1,294 |
0,048 |
12 |
|
P.leo |
0,885 |
0,441 |
11 |
Однако в плане нашей темы интереснее другое. Эксперименты показали, что аллометрический экспонент очень строго фиксирован наследственно. Наследственные изменения а (изменения пропорций черепа) возможны только при резком изменении условий существования животных, влияющих на скорость их роста [Шварц, 1961]. Поэтому совершенно очевидно, что если бы наблюдающиеся изменения а определялись преимущественно не наследственными механизмами, то особенно резкие различия всегда обнаруживались бы между весенними и осенними популяциями, так как от весны к осени происходит смена поколений животных, ро-
{173}
дившихся и выросших в совершенно разных условиях. От осени к весне подобных изменений не происходит. В это время полевки не размножаются, следовательно, это те же животные, но состав их изменился за счет отмирания определенной группы особей. Поэтому все случаи, когда изменения от осени к весне более значительны, чем от весны к осени, совершенно неоспоримо свидетельствуют о дифференциальной смертности, которая приводит к изменению генетического состава популяции. Приведенные в табл. 17 данные показывают, что изменения этого типа нередко проявляются очень резко.
У нас нет никаких оснований полагать, что рассматриваемый признак, отражающий изменения в соотносительной скорости роста различных частей черепа и черепа в целом, в рассматриваемом положении уникален. Наоборот, можно быть уверенным, что точно такие же данные были бы получены и при анализе других признаков, более существенных с экологической точки зрения. Об этом свидетельствуют многочисленные примеры изменений средней нормы изменчивости популяции по годам, которые не могут быть полностью объяснены непосредственной реакцией животных на конкретные условия среды [Тимофеев-Ресовский, 1940, 1964; Шмальгаузен, 1946; Шварц, 1959].
Таким образом, мы приходим к выводу, что морфофизиологическая специфика возрастных и сезонных генераций определяется не только специфическими условиями их развития, но и преобразованием генетической структуры популяций. Колебания «качества» популяции — столь же характерное ее свойство, как и колебания численности. Явления эти взаимно связаны: изменения численности (в особенности резкие) сопровождаются изменением генетического состава популяции. Это приводит нас к важным теоретическим выводам, составляющим основную часть этой главы.
Изменение возрастной структуры популяции сопровождается изменением ее генетической структуры. Как указывалось, об этом свидетельствуют многочисленные литературные данные и некоторые наши материалы. Это значит, что если по каким-то ни было причинам смертность животных разного возраста будет существенно различной, то это приведет к существенному изменению генетической структуры популяции со всеми вытекающими отсюда последствиями. Назовем этот процесс условно возрастным отбором. Нетрудно представить себе, каков механизм взаимосвязи между возрастной структурой популяции и изменением ее генетического состава. Воспользуемся гипотетическим примером отбора грызунов на «зимостойкость».
В течение зимы происходит сдвиг генетической структуры популяции в сторону «зимостойких» особей. (Какими физиологическими причинами эта «зимостойкость» определяется, для нас в данном случае не важно, но в отдельных случаях об этом можно было бы сказать и кое-что конкретное.) Поэтому в группе «ста-
{174}
риков» подобные «зимостойкие» особи будут относительно более многочисленными, чем в группе молодых. Логично полагать, что особенно резко это будет выражено в более суровые зимы, когда преимущество «зимостойких» особей проявится сильнее. В этих условиях индивидуальный отбор будет работать в пользу «зимостойких» особей, но возрастной отбор должен действовать в обратном направлении, так как в суровые зимы отмирание «стариков» выражено особенно резко. Конечный эффект будет зависеть от соотношения сил этих двух форм отбора. Важно, однако, что они могут быть противоположно направленными и что возможны ситуации, когда именно мягкая зима поведет к резкому повышению в популяции животных зимостойкого типа. Это результат не той формы отбора, которой придается наибольшее (если не исключительное) значение, а результат изменения возрастной структуры популяции. Экспериментальное исследование различных сторон этого вопроса, к которому мы только еще приступаем, может представлять большой интерес не только для теории (в частности, для теории микроэволюции), но и для некоторых отраслей практики, о чем будет сказано ниже. Важно подчеркнуть: теоретический анализ приводит нас к выводу, что изменение экологической (в данном случае — возрастной) структуры популяций неразрывно связано с изменением ее генетической структуры со всеми вытекающими отсюда последствиями. Изменение экологической структуры популяций должно, следовательно, рассматриваться в качестве важнейшего фактора микроэволюционного процесса.
Приведенные факты показывают, что сезонное изменение генетической структуры популяции — широко распространенное, если не всеобщее, явление. У многих видов животных оно проявляется в генетическом своеобразии разных генераций. Отсюда следует, что изменение возрастной структуры популяции с неизбежностью приводит к изменению общего генетического состава популяции и более быстрой ее эволюции, чем под влиянием индивидуального естественного отбора. В относительно стабильных условиях изменение генетической структуры популяции, связанное с динамикой ее возрастного состава, имеет характер осцилляций около некоторой многолетней средней (аналогия с колебаниями численности). При изменении условий среды возрастной отбор может явиться фактором быстрых эволюционных преобразований. Изменение климата может, например, привести к выпадению отдельных генераций и соответствующему изменению ее генетической структуры, которая не будет уже восстановлена в последующие годы. Как показали приведенные выше примеры, генетические различия между генерациями могут превосходить средние различия между популяциями, в течение тысяч поколений развивавшихся самостоятельно, но нетрудно себе представить, какое значение может иметь феномен «возрастного отбора» для темпов микроэволюции. Нельзя, однако, забывать, что «возрастной
{175}
отбор» работает на фоне индивидуального. Именно последний создает генетические различия между генерациями. То, что индивидуальный отбор при изменении генетической структуры сезонных генераций работает с большей эффективностью, чем при дифференциации популяций, хорошо понятно, так как условия развития, например, летних генераций животных в степи и тундре (берем крайний пример) более сходны, чем условия развития тундровых животных ранней весной и поздним летом. Если для сравнения воспользоваться популяциями из более близких ландшафтных зон, то хорологические и хронологические различия в условиях развития животных выявляются с полной отчетливостью. Возрастной отбор (в указанном выше понимании) создает условия для мобилизации тех изменений, которые возникают в популяциях под воздействием индивидуального отбора. Их совместное действие в конечном итоге определяет темпы преобразования популяций. При длительном (или необратимом) изменении климата это преобразование закрепляется и ведет к эволюционным изменениям. Однако и временное изменение генетической структуры популяции может иметь большое эволюционное значение при изменении пространственной структуры популяции.
Для того чтобы составить представление о том, насколько широко распространено явление возрастного отбора, необходимо учитывать, во-первых, возможный размах «обычной» динамики возрастной структуры популяций и, во-вторых, физиологические отличия между животными разного возраста.
Первый вопрос настолько хорошо изучен, что здесь достаточно привести две таблицы, характеризующие динамику возрастного состава животного, обладающего большой продолжительностью жизни (песец, табл. 19), и животных-эфемеров (полевки, табл. 20). Таблицы эти вряд ли нужно комментировать. Они ясно свидетельствуют о масштабах динамики возрастной структуры популяции. Аналогичные данные получены в настоящее время для многих видов копытных [Скалон, 1960; Pimlott, 1961], ластоногих [Laws, 1956], хищников [Wood, 1959], грызунов [Лаврова, Карасева, 1956; Поляков, 1964], птиц [Pinowski, 1965] и других животных. Исследования в этом направлении развиваются очень энергично, и накопление фактического материала происходит быстро. Важно поэтому обратить внимание, что возрастная структура популяции отдельных видов нередко определяется влиянием хищников [Лаврова, Карасева, 1956; Базиев, 1967; Knight-Johnes, Moyse, 1961 и др.]. Отсюда следует, что изменение численности одного вида влияет не только на численность другого, но и на структуру его популяций, а следовательно, и на генетический состав. Если различия в генетической структуре разных возрастных групп существенны, то в ситуациях, описанных выше, они не могут не повлиять на динамику генетической структуры популяции в целом.
Второй вопрос требует экологической оценки тех физиологических различий, которые обнаруживаются при сравнении живот-
{176}
Таблица 19
Численность песца к началу промыслового сезона в Ямало-Ненецком национальном округе в разные годы
[по В. С. Смирнову]
|
Показатель |
1955 |
1956 |
1957 |
1958 |
|
Общая численность |
42861 |
35893 |
54895 |
34025 |
|
Число молодых в данной генерации |
31082 |
27409 |
43334 |
6936 |
|
Процент молодых |
72,5 |
76,4 |
79 |
20,4 |
|
Число молодых на пару взрослых |
5,28 |
6,46 |
7,51 |
0,51 |
|
Показатель |
1959 |
1960 |
1961 |
1962 * |
|
Общая численность |
26094 |
37480 |
41743 |
19026 |
|
Число молодых в данной генерации |
18961 |
29886 |
28453 |
259 |
|
Процент молодых |
67,4 |
79,8 |
68,2 |
1,4 |
|
Число молодых на пару взрослых |
5,32 |
7,88 |
4,28 |
0,03 |
* Данные за 1962 г. определены с невысокой точностью.
Таблица 20
Зависимость возрастной структуры популяций полевок от погодных условий
(южное Зауралье) *
|
Вид |
Количество молодых, % к общему числу добытых |
Примечание |
|
|
1950 |
1951 |
||
|
Зодяная полевка |
1,2 |
22 |
В июне 1950 г. только 13% молодых самок водяных полевок принимало участие в размножении. В 1951 г. в это же время размножалось 57% молодых самок, а 21% принимал участие в размножении дважды |
|
Полевка-экономка |
Нет |
2,8 |
|
|
Узкочерепная полевка |
12,0 |
23,0 |
|
|
Красная полевка |
Нет |
9,0 |
|
* 1950 г.— весна ранняя, снег полностью сошел в конце апреля, 1951 г.— весна ранняя, снег полностью сошел в начале апреля. Указан процент прибылых особей во второй половине мая.
{177}
ных разного возраста. Так как решение этого вопроса создает предпосылки для оценки вероятной распространенности возрастного отбора, остановимся на нем с известной подробностью.
Хорошо известно, что многие факторы внешней среды, существенно не отражающиеся на жизни взрослых животных, ведут к гибели молодняка. Пересыхание водоемов ведет к массовой гибели личинок амфибий, но существенно не отражается на численности взрослых особей, близкой к нормальной. Весенние паводки и осенние ливни особенно губительно отражаются на беспомощном молодняке млекопитающих. Подобные факты хорошо известны и для разных видов описаны с необходимой детализацией. Они показывают, что селективная смертность—явление обычное. Однако различие в смертности молодых и взрослых в интересующем нас здесь эволюционном плане представляет относительно меньший интерес, так как генетическая структура молодняка до перехода к самостоятельному образу жизни должна быть весьма близка к генетической структуре их родителей. Вероятно, большее значение имеет различие в смертности животных разного возраста, но уже перешедших к самостоятельному образу жизни. По этому вопросу данных значительно меньше, но физиологические различия между разновозрастными группами животных столь значительны, что они должны вызывать селективную смертность.
Несмотря на отсутствие существенных возрастных изменений в способности протоплазмы к потреблению кислорода, по крайней мере в тех возрастных пределах, которые нас здесь интересуют, основной обмен в силу уменьшения числа функционирующих клеток и ряда других причин с возрастом падает. В соответствии с этим падает и способность к максимальному потреблению кислорода, что имеет особое значение при повышении физической нагрузки.
Это падение становится заметным уже в относительно молодом возрасте. У человека в 40 лет максимальное потребление кислорода на 20% меньше, чем в 25. В среднем возрасте наблюдаются существенные изменения и ряда других физиологических функций: уменьшается насыщение кислородом артериальной крови, повышается кровяное давление, возрастает содержание молочной кислоты и понижается щелочной резерв крови (это особенно хорошо заметно в условиях, повышенной физической нагрузки), падает эластичность кровеносных сосудов, снижается максимальный пульс, падает способность кишечника к всасыванию некоторых элементов, необходимых для нормального функционирования организма. С возрастом падает не только интенсивность метаболизма организма в целом, но и метаболическая активность тканей и клеток. Трудно сказать, с какого возраста (у разных видов, конечно, различного) этот процесс принимает ощутимые размеры. Во всяком случае согласованность данных, полученных различными исследователями при помощи различных методов, показывает, что у крыс, например, к двум годам это падение до-
{178}
стигает весьма существенных размеров даже при сравнении их с вполне взрослыми животными в возрасте более года [Weinbach, Garbus, 1956; Barrows et al., 1957].
Особенно важно подчеркнуть, что с возрастом изменяются те физиологические особенности организма, которые определяют его реактивность в ответ на неблагоприятные или просто изменяющиеся условия внешней среды. С возрастом нарушается центральная нервная корреляция и падает скорость нервных импульсов. Происходит функциональное изменение деятельности эндокринной системы, которое отражается в прогрессивном уменьшении размеров клеточных ядер желез внутренней секреции, в падении митотической активности, разрастании соединительной ткани и как следствие этого в падении выделения некоторых гормонов [Symposium Institute of Biology, 1956]. Животные старшего возраста обладают пониженной терморегуляторной способностью и относительно меньшей способностью создавать физиологические резервы. Все это не может не приводить и действительно приводит к существенным различиям у животных различного возраста в реакциях на изменение внешних условий.
Эту закономерность очень удобно проиллюстрировать отношением разновозрастных животных к трем важнейшим факторам внешней среды: температуре, кислородному режиму и питанию.
Рядом исследователей констатировано резкое падение у животных старшего возраста способности приспособления к понижению температуры. Показано [Grad, Krai, 1957], например, что смертность мышей линий С57В в возрасте 16–22 месяцев много выше, чем у 4–9-месячных животных. С возрастом снижается и способность крыс адаптироваться к низким температурам. В опытах авторов 60% «адаптированных» старших крыс погибло в течение недели при температуре, при которой ни одна из «адаптированных» молодых не погибла.
Аналогичные результаты получены при сравнении разновозрастных животных по их реакции на снижение концентрации кислорода в атмосфере. Установлено, например, что морские свинки весом около 300 г значительно устойчивее к недостатку кислорода, чем животные старшего возраста весом около 500 г. Различная чувствительность животных разного возраста к качеству кормов иллюстрируется повышением потребности старших животных к витамину B1, являющемуся одним из важных катализаторов окислительно-восстановительных систем клеток [обзор данных см. Шварц, 1960].
Имеются данные, позволяющие полагать, что смертность старших возрастных групп превышает смертность молодых животных. Это было, например, в самой общей форме показано на лесной мыши в работе с применением весьма совершенной методики биометрической обработки материала [Hacker, Pearson, 1944]. Некоторыми авторами отмечается дифференцированное истребление хищниками разновозрастных животных [Фолитарек, 1948; Лав-
{179}
рова, Карасева, 1956]. Есть основания полагать, что восприимчивость молодых животных к болезням выше, чем у старых [Поляков, Пегельман, 1950]. Это, по-видимому, справедливо и в отношении некоторых гельминтозов. Так, например, в отдельных местностях Западной Европы кокцидиозу подвержены почти исключительно молодые зайцы.
Особый интерес представляют данные, показывающие, что в основе дифференцированной смертности может лежать различное отношение животных разных возрастов к среде обитания.
И. Я. Поляков и С. Г. Пегельман [1950] показали, что при температуре 35°, когда половозрелые общественные полевки гибнут, более молодые животные энергично растут и заметно не теряют жизнеспособности. В соответствии с этим во время обычных в Азербайджане засух происходит интенсивное отмирание старших возрастов и общее «омоложение» популяции.
Однако неблагоприятное сочетание зимних условий животные старших возрастов переносят легче [Поляков, 1956].
Совершенно естественно, что биологические отличия между животными разного возраста с неизбежностью вызывают их различную смертность. К сожалению, данные, которыми располагает экология по этому вопросу, все еще скромны. Если зависимость между возрастной и генетической структурой популяции будет установлена, а причины селективной смертности животных выяснены с необходимой точностью, то тем самым будет создана возможность не только для предвидения генетических последствий определенного сочетания внешних условий, но и для вмешательства в начальные этапы микроэволюционного процесса. Нам представляется, что исследование этих процессов — одна из наиболее актуальных задач эволюционной экологии.
В нашем исследовании, проведенном совместно с В. Г. Ищенко, в качестве объекта была избрана остромордая лягушка (Rana arvalis), в популяциях которой почти по всему ареалу встречаются два генетических варианта — striata и maculata. Striata имеют хорошо заметную дорсальную полосу, maculata этой полосы не имеют, для них характерна пятнистая окраска спины. Исследования проводились на Южном Урале, в Ильменском заповеднике, в течение двух лет, на одном и том же месте, в одно время (начало августа). Обследовалась относительно изолированная популяция лягушек на небольшом болоте, прилегающем к озеру Миассово. Подразделение лягушек на возрастные группы основывалось на анализе кривых распределения длины тела. Этот метод не гарантирует от ошибок, но при работе с массовым материалом дает результаты вполне удовлетворительной точности. Полученные данные представлены в табл. 21. Их анализ приводит к следующим выводам.
В 1966 г. во всех возрастных группах striata составляли немногим менее 50%. Различия в генетическом составе животных разных лет «рождения» недостоверны. Иная ситуация сложилась
{180}
Таблица 21
Изменение относительной частоты встречаемости варианта striata (в %) в популяции Rana arvalis по годам
(Ильменский заповедник, Челябинская область)
|
Возрастная группа |
1966 г. |
1967 г. |
|
0+ (сеголетки) |
39,1±2,43 (402) * |
28,6±6,04 (56) |
|
1+ |
49,2±4,35 (132) |
42,6±6,33 (61) |
|
2+ |
44,1±3,07 (261) |
43,3±4,04 (150) |
|
3+ |
44,4±3,70 (180) |
37,1±3,31 (213) |
|
4+ |
42,1±8,1 (38) |
61,1±5,74 (72) |
|
5+ |
100(1) |
64,9±7,84 (37) |
* В скобках — число обследованных особей
в 1967 г. В группе сеголеток striata достоверно меньше, чем в старших возрастных группах (критерий статистической достоверности t=2,07). Наоборот, среди лягушек старшего возраста striata резко преобладают. Различие между возрастной группой 3+ и старшими статистически абсолютно достоверно (t=3,05). Уже эти данные представляют интерес. Они еще раз подтверждают положение о том, что генетический состав различных генераций животных одной популяции различен, и это различие существенно. С другой стороны, эти же данные показывают, что генетический состав отдельных генераций не остается постоянным, он изменяется, притом вполне закономерно. На нашем материале это отчетливо проявляется при сравнении лягушек старшего возраста. В 1966 г. среди лягушек в возрасте 3+ striata составляли 44,4%, в 1967 г. в этой генерации (сейчас уже в возрасте 4+) они составляли большинство (t=2,29). В том же направлении изменился и генетический состав генерации предшествующего года (сравниваются четырехлетки в 1966 г. с пятилетними в 1967 г.; t=2,24).
Эти данные показывают, что интенсивность отбора в природных условиях весьма значительна, неизмеримо больше, чем его эффективность, о которой мы судим на основе сдвига средней нормы изменчивости популяции в целом. Причина заключается не только в закономерном изменении направления отбора при изменении условий среды, которое вызывает гомеостатическое колебание генетического состава популяции, но и в разной реакции животных разного возраста па аналогичные изменения условий существования. Представленный материал позволяет утверждать, что относительная смертность striata и maculata в разных возрастных группах различна. В старших возрастных группах maculata характеризуются более высокой смертностью (возможно, что только в тех условиях, которые сложились в годы нашей работы),
{181}
поэтому в группе лягушек старше четырех лет значительно преобладает striata.
В настоящее время мы не можем привести полный генетический анализ нашего материала, так как у нас нет данных, характеризующих смертность гетерозигот. Однако для анализа значения динамики возрастной структуры популяции в микроэволюционном процессе мы располагаем достаточной информацией. Воспользуемся в данном случае хорошо известным приемом мысленного эксперимента. Допустим, что в силу каких-то причин младшие возрастные группы лягушек вымерли и популяция начинает восстанавливаться за счет животных возраста 5+. Это значит, что ядро производителей почти на 70% будет состоять из striata, что более чем на 20% превышает среднее (по всем генерациям) количество этой формы в популяции 1966 г. Подобный сдвиг в генетической структуре популяции мог бы произойти под воздействием индивидуального отбора даже высокой интенсивности лишь за много лет. Как будет подробно показано ниже, мы не противопоставляем индивидуальный отбор возрастному. Возрастной отбор (изменение возрастной структуры популяции) мобилизует генетические потенции, создаваемые индивидуальным отбором, и в силу этого многократно увеличивает его эффективность. Можно сказать, что возрастной отбор снимает противоречие между интенсивностью и эффективностью естественного отбора.
Полученные нами данные показывают также, что резкие изменения возрастной структуры полиморфной популяции, подобные тем, которые мы приняли в нашем мысленном эксперименте,— реальность. Достаточно обратить внимание на резкое снижение относительной доли сеголеток в 1967 г. (по сравнению с 1966 г.) и не менее резкое увеличение числа наиболее старых животных. Изменение возрастной структуры популяции может быть вызвано не только различным числом молодых животных, успешно закончивших метаморфоз (в 1967 г. большое число головастиков погибло в результате засухи и сопутствующих явлений), но и изменением условий среды на разных участках территории, занятой популяцией. Наблюдения показали, что лягушки разного возраста населяют разные участки. Поэтому любое локальное изменение условий среды (засуха, ранняя гололедица и т. и.) может вызвать резкое изменение возрастной структуры популяции, не говоря уже о возможной дифференциации смертности, связанной с физиологическими особенностями животных разного возраста. Резкая перестройка генетического состава популяции оказывается при этом неизбежной.
Естественно, что изменение роли различных возрастных групп в поддержании численности популяции определяется не только селективной смертностью, но и изменением характера размножения.
Зависимость интенсивности размножения от различного сочетания внешних условий — одна из наиболее полно разработанных
{182}
глав экологии. Поэтому нам нет нужды приводить примеры зависимости изменения структуры популяций животных разных видов от количества рекрутируемого в популяцию молодняка. Однако связь интенсивности размножения с генетической структурой популяции до сих пор не изучена, и это, вероятно, не дает возможности в полной мере оценить значение экологических механизмов начальных стадий дивергенции популяций.
Необходимо иметь в виду, что интенсификация размножения даже с чисто экологической точки зрения (динамика численности) не представляет собой простого увеличения относительного обилия в популяции молодых животных. Эколого-эндокринологические исследования ясно показали, что резкая интенсификация размножения находится в тесной связи с плотностью популяций [Christian, 1961; 1963; Wynne-Edwards, 1962 и др.]. После снижения численности не только увеличивается плодовитость и число самок, участвующих в размножении, но и увеличивается скорость полового созревания молодых животных. Зависимость интенсивности размножения от плотности популяций лучше всего изучена на млекопитающих, но имеются наблюдения, показывающие, что она проявляется у других позвоночных [Fehringer, 1962] и у насекомых [Pajunen, 1966]. Литература по этому вопросу уже сейчас значительна. Накопленные данные ясно показывают, что связанное с изменением плотности изменение интенсивности размножения приводит к резкому изменению возрастной структуры популяций. Очень важно, что конкретное проявление этой закономерности даже у очень близких видов может быть принципиально различным [Lidicker, 1965].
В некоторых случаях изменение возрастного состава размножающихся животных выходит за рамки нормальной экологической характеристики вида. Мы ограничимся лишь одним примером. Как известие, бурозубка в средних и южных широтах обычно в год своего рождения в размножение не вступает. Однако Штейн [Stein, 1961] показал, что число самок, вступающих в размножение в год своего рождения, зависит от плотности популяции. Если в обычные годы размножающиеся сеголетки составляют 1–2%, то после резкого спада их количества это число у малой бурозуб-ки и куторы доходит до 35 %! Значение подобных явлений в динамике численности животных оценено уже давно и хорошо понятно, но их значение в динамике качества популяции еще никто оценить не пытался. Между тем ясно, что коль скоро в течение года происходит перестройка генетической структуры популяции, что разные поколения генетически не тождественны, то «незаконное» вступление в размножение молодых землероек не может не вызвать нарушения типичной для вида цикличности генетического состава популяции. Примеры, приведенные ранее, показывают, что и в данном случае мы сталкиваемся с новым и очень интересным разделом эволюционной экологии.
Возрастной отбор не только объясняет возможные причины из-
{183}
менения скорости эволюции, но и предпосылки для создания теории управления качественным составом популяций. Все факторы, изменяющие возрастную структуру популяции, автоматически приводят к изменению ее генетической структуры. Если зависимость между экологической и генетической структурой популяции известна, то разработка методов управления генетическими преобразованиями популяций сталкивается лишь с техническими, а не принципиальными трудностями.
Развиваемые нами взгляды на значение экологической структуры популяции в микроэволюционном процессе заставляют с новой точки зрения смотреть и на значение так называемой неизбирательной элиминации.
О ЗНАЧЕНИИ НЕИЗБИРАТЕЛЬНОЙ ЭЛИМИНАЦИИ
Господствующая в настоящее время теория эволюции принимает следующие основные постулаты:
1. Творческую роль играет только индивидуальный естественный отбор внутри популяции.
2. Особо интенсивная элиминация лишь в исключительных случаях может иметь избирательный характер.
3. Чем более активны истребительные факторы среды, тем более общий характер принимает элиминация, теряющая перед лицом стихийных сил природы свой избирательный характер.
4. Мощные факторы среды, ведущие к резкому снижению численности животных, не могут вызвать направленных изменений в структуре популяций; возникающие изменения случайны.
Была сделана попытка на основе математического моделирования определить скорость генетических преобразований популяций. Полученные выводы сводятся к следующему. На начальных стадиях направленных генетических преобразований популяций для достижения видимого эффекта требуются сотни тысяч поколений, затем преобразования идут быстрее, но и в этом случае период заметных сдвигов измеряется тысячами поколений.
Попытаемся проанализировать эти положения с учетом данных о генетической специфике сезонных и возрастных генераций животных. Подчеркнем один из основных постулатов теории генетико-автоматических процессов: неизбирательная элиминация творчески бессильна, она создает лишь случайные изменения генетической структуры популяции со всеми вытекающими отсюда последствиями (глава II). Так ли это? Ведь неизбирательная элиминация всегда происходит в какое-то определенное время года и на фоне определенной возрастной структуры и сезонного изменения генетической структуры, вызываемого избирательным отбором. Если элиминация неизбирательна, то это значит, что представленные в популяции генотипы элиминируются в соответствии с их относительным количеством. Это и создает впечатление, что
{184}
никаких направленных изменений неизбирательная элиминация создать не может.
Попытаемся, однако, проанализировать этот вопрос более глубоко. Допустим, что мы имеем дело с диморфной популяцией» представленной генетическими вариантами (фазами) А и В. Фаза А в летнее время интенсивнее размножается, и соответственно от весны к осени происходит относительное увеличение ее численности. Реальность подобного предположения могла бы быть подкреплена большим числом фактов.
Допустим, что весной структура популяции выглядит так: 50А+150В (абсолютное выражение коэффициентов значения не имеет, оно призвано лишь отражать относительное обилие в популяции разных генетических вариантов). За период размножения А увеличилось в 10 раз, В — в 2 раза. Тогда осенью генетическая структура популяции будет иметь вид: 500А+300В.
Посмотрим, к чему приведет неизбирательная элиминация, действующая в разное время года. Допустим, что в результате элиминации численность животных уменьшилась в 50 раз. Так как элиминация неизбирательна, то каждая фаза снижается в численности в равной степени (с учетом статистических погрешностей). Весной — (50А+150В): 50=1А+3В. Вероятность полного исчезновения А больше, чем В, а при повторном элиминировании вымирание гомозиготных А практически неизбежно. Осенью — (500А+300В): 50=10А+6В. В этом случае вероятность исчезновения В больше, чем А. Уже эта простейшая модель показывает, что так как неизбирательная элиминация проходит на фоне закономерных сезонных колебаний генетической структуры популяции, то в конечном итоге она направленно изменяет структуру популяции в целом. Естественно, что в природе все происходит значительно сложнее: популяция представлена не двумя генотипами, а множеством, различия в потенциале их воспроизводства, вероятно, менее значительны, при очень низкой численности отдельных вариантов вероятность их гибели уменьшается, но общая закономерность выражается нашей моделью правильно. Она построена на точно доказанных закономерностях. О том, что генетическая структура популяции подвержена сезонной изменчивости, мы уже говорили. О том, что «неизбирательная элиминация» почти всегда имеет характер сезонного бедствия, пожалуй, и говорить не стоит, это слишком хорошо известно. Возврат холодов, заморозки, паводки, ливни, эпизоотии — это все сезонные явления, да притом для разных видов преимущественное значение имеют одни из них, другие — второстепенное. Трудно, пожалуй, назвать хотя бы одну форму неизбирательной элиминации (за исключением землетрясений и вулканических извержений), которая по своей природе не была бы сезонной. Отсюда следует, что, по крайней мере в очень многих случаях, неизбирательная элиминация может оказать на развитие популяции направленное действие.
Сказанное, конечно, не означает ни отрицания, ни умаления
{185}
выдающегося значения исследований по популяционной генетике, но в настоящее время они не могут ограничиваться чисто теоретическими или лабораторными. Они должны исходить из реальных представлений по экологии популяций. Попытаемся поэтому приблизить нашу модель к природе.
Допустим, что фаза А отличается от В несколько большей скоростью полового созревания. В соответствии с этим животные этой фазы за нормальный сезон размножения успевают дать 2 помета, животные второй фазы — только один. Плодовитость животных обеих фаз и их смертность в течение летнего периода одинаковы. В таком случае нормальный жизненный цикл популяции будет выглядеть так: весна—50А+150В (1:3); осень — 50А (производители)+150А (первый помет)+150А (второй помет)+150В (производители) +450В (первый помет). Численность А увеличивалась за сезон размножения в 7 раз (с 50 до 350), В — в 4 раза (со 150 до 600). Для того чтобы генетический состав популяции оставался постоянным, необходимо допустить, что в зимнее время имеет место дифференцированная смертность: численность А снижается в 7 раз, численность В — в 4 раза. Осень — 350А+600В (7: 12), весна-50А+150В (1:3).
Допустим теперь, что ранней осенью имеет место неизбирательная элиминация, численность животных резко снижается и при этом полностью гибнет весь второй помет. Это также вполне реальное допущение: при ранних и сильных заморозках, например, происходит неизбирательная элиминация животных, ведущих самостоятельный образ жизни, но несамостоятельный молодняк гибнет полностью. В этих условиях преимущество генетического варианта А проявиться не может, и динамика генетической структуры популяции резко изменяется: весна — 50А+150В, осень — 50А+150А+150В+450В=200А+600В (напоминаем, что коэффициенты указывают лишь на соотношение разных форм в популяции, а не на их абсолютное количество).
Зимой сокращение численности происходит по обычной схеме: А уменьшается, получаем весенний состав популяции: 30А+150В (1:5).
Допустим, что и следующей осенью произойдет элиминация с такими же сопутствующими явлениями. Тогда к концу сезона размножения имеем: 30А+90А+90А+150В+450В=120А+600В. Соответственно следующей весной имеем: 17А+150В. При повторении сходной ситуации в третий раз генетический состав нашей популяции примет вид: 6А+150В. Если теперь ситуация, вызывающая элиминацию, произойдет до начала размножения и снизит общую численность популяции в 10 раз, то по теории вероятности генетический вариант А вообще исчезнет из популяции.
В реальной природной обстановке А не исчезнет, а сохранится в популяции в гетерозиготном состоянии. Наш пример показывает, однако, насколько быстро может произойти направленная генетическая перестройка популяции под влиянием ненаправленного
{186}
фактора среды (неизбирательная элиминация). В рамках нашей темы нам особенно важно подчеркнуть, что непосредственной причиной изменения генетической структуры популяции является изменение ее экологической (в нашем примере — возрастной) структуры.
Введем в нашу модель еще одно экологическое уточнение: при снижении плотности популяции плодовитость животных возрастает. Допустим, что ранней весной (до начала размножения) произошла неизбирательная элиминация. Соотношение генетических вариантов не изменилось (50А+150В), но общая численность животных снизилась. В соответствии с действием «факторов, зависящих от плотности» (density dependent factors), плодовитость животных увеличилась, и осенью на каждую пару взрослых приходится не 6, а 12 молодых. Тогда осенняя структура популяции может быть выражена так: 50А+300А+300А+150В+900В=650А+1050В (13:21). И в данном случае изменение генетической структуры популяции было вызвано изменением ее экологической структуры.
Попытаемся еще более приблизить нашу модель к реальной экологической обстановке. Действие «факторов, зависящих от плотности», с особой силой проявляется непосредственно после разрежения популяции. Поэтому в нашем примере резко увеличенным должен быть первый помет. Второй помет будет уже менее многочисленным, так как плотность популяции после пополнения ее сеголетками повышается (не говоря уже о закономерном снижении плодовитости к осени). Поэтому осенняя популяция должна принять вид: 50А+300А (первый помет)+150А (второй помет)+150В+900В (первый помет) =500А+ 1050В (10:21). Структура популяции изменилась еще более существенно.
Рассматриваемый здесь вопрос еще не привлек к себе пристального внимания экологов. Поэтому представляется важным отметить, что в пользу развиваемых нами взглядов свидетельствуют общие теоретические соображения, основанные не только на твердо установленных экологических закономерностях, но и на некоторых (пока еще немногочисленных) прямых наблюдениях, показывающих, что резкое изменение численности сопровождается не случайным, а закономерным изменением генетического состава популяций. В этом отношении особый интерес представляет работа Б. К. Павлова [1965], который по нашему совету изучал динамику полиморфизма белок и сумел установить связь изменений частоты встречаемости отдельных генетических вариантов с их морфофизиологическими особенностями. Белки Восточной Сибири представлены несколькими цветовыми вариациями: краснохвостки, чернохвостой и промежуточная группа — бурохвостки. Каждой популяции свойственно свое отношение цветовых вариаций. В южных горно-таежных лесах с преобладанием кедра сибирского доминируют чернохвостые особи (до 80%), краснохвостки составляют 5–6%, бурохвостки — 10–15%, в северных лист-
{187}
венничных лесах соответственно 30–40%, 18–20%, 40–50%. На территории, занимаемой одной популяцией, черные белки заселяют темнохвойные леса, светлые — светлохвойные.
Характерное для каждой популяции соотношение цветовых вариаций не остается постоянным, а изменяется по годам. Общая закономерность такова, что при высокой численности преобладают чернохвостые белки, после депрессии численности увеличивается относительная многочисленность бурохвостых. В определенные годы количество чернохвостых достигает 50% даже в северных «светлых» популяциях.
Особи различных цветовых вариаций отличаются рядом морфофизиологических показателей. Краснохвостки легче переносят недостаток основных кормов, длина кишечника у них больше, чем у чернохвосток и бурохвосток. Вследствие этого они обладают большей способностью использовать грубые растительные корма. В северных популяциях относительный вес печени у краснохвосток больше, чем у чернохвосток. В южных популяциях эти различия недостоверны. Относительный вес надпочечников у бурохвосток значительно ниже, чем у краснохвосток и чернохвосток. При оптимальной плотности краснохвостые и чернохвостые белки по относительному весу надпочечников не отличаются. При резком ухудшении условий существования различные цветовые вариации отличаются по плодовитости, при этом у краснохвосток она выше. Различные цветовые вариации обладают различными биологическими особенностями.
Можно предполагать, что соотношение различных цветовых вариаций находится под контролем естественного отбора. Географическая изменчивость фенотипической структуры популяции может служить некоторым доказательством в пользу этого положения. Для популяций с неустойчивым типом динамики численности свойственна структура с преобладанием бурохвостых и краснохвостых особей. Для популяций с устойчивым типом динамики численности свойственно преобладание чернохвостых особей (количество бурохвостых особей крайне незначительно, краснохвостых почти всегда около 5–6%).
Особый интерес представляют данные Б. К. Павлова, обобщенные им в переданной мне рукописи 1967 г., которые характеризуют генетический состав разных возрастных групп белок. В Тогодинской популяции в 1962 г. в разных возрастных группах относительное обилие цветовых вариантов выражалось следующими цифрами (указан процент белок в следующем порядке: чернохвостые, бурохвостые, краснохвостые).
Сеголетки 85,5±2,9 12,4±2,7 2,2±1,2
1 год 65,8±4,1 27,6±4,5 6,6±2,5
2 года 14,8±9,9 71,4±12,5 14,8±9,9
Эти данные делают очевидным, что если по каким-либо причинам возрастная структура популяции изменится, то это повлечет за со-
{188}
бой резкое изменение генетической структуры популяции в целом. Для того чтобы представить себе конкретный ход этих изменений, необходимо знать характер детерминированности рассматриваемых генетических вариантов (в настоящее время не известны, но в их неизбежности трудно сомневаться). Возрастной отбор может привести к быстрой перестройке популяций. Исследования Б. К. Павлова делают это бесспорным.
Весьма существенно, что изменение генетической структуры популяций происходит и под влиянием промысла, причем на животных разного возраста промысел оказывает разное воздействие.
Таблица 22
Изменение аллометрического экспонента а в экспериментальных популяциях белок
(по данным Б. К. Павлова) *
|
Период обследования |
Ширина межглазничного промежутка |
Длина зубного ряда |
Ширина между верхними коренными зубами |
|||
|
самцы |
самки |
самцы |
самки |
самцы |
самки |
|
|
До промысла |
-0,066 ±0,019 |
+0,265 ±0,0019 |
-0,022 ±0,017 |
- -0,252 ±0,012 |
+0,002 ±0,25 |
-0,386 ±0,06 |
|
После промысла |
+0,926 ± 0,085 |
-0,175 ±0,021 |
+0,375 ±0,014 |
-0,165 ±0,015 |
+0,531 ±0,09 |
+0,151±0,002 |
* Все показатели по отношению к кондилобазальной длине черепа.
Как указывалось ранее, изменение генетической структуры популяций проявляется в изменении аллометрического экспонента — признака, характеризующего генетический состав популяции в целом. Б. К. Павлов провел специальный опыт на экспериментальной площадке, который показал, что за 10 дней промысла генетический состав популяции белок изменился. Аллометрический экспонент основных показателей за этот период изменился очень существенно (табл. 22). На контрольных площадках изменений отмечено не было. Перемещений белок по территории не происходило.
Изменение генетической структуры популяции Б. К. Павлов изучал и на основе использования в качестве генетического маркера наличия межтеменной кости (анализ значения подобных признаков см. [Berry, 1963]). До промысла белки с этим признаком составляли 14,9% в популяции, после промысла — 23,8% (различия статистически достоверны). Особенно резкие изменения произошли в группе молодых белок: до промысла — 3,2%, после промысла — 18,2% (t=6,7). Достоверность этих данных Б. К. Пав-
{189}
лов подтверждает не только статистической обработкой материала, но и поголовным отстрелом всех белок на экспериментальной площадке. Анализируя свой материал, он отмечает, что в некоторых случаях избирательное действие промысла совпадает с направлением естественного отбора. Преобразования популяции могут произойти весьма быстро.
Трудно сказать, чем вызвана избирательность промысла. Несомненно, генетические маркеры связаны с какими-то особенностями биологии белок, определяющими их поведение. Изменение аллометрического экспонента наводит на мысль о том, что эти особенности определяются разной активностью животных, так как изменение может быть связанным с изменением скорости роста животных [Шварц, 1961]. Важен, однако, несомненный факт, доказанный работами Б. К. Павлова: динамика численности, определяемая как естественными причинами, так и промыслом, вызывает изменения генетической структуры популяций, которые происходят особенно резко в том случае, когда вступает в силу возрастной отбор (в упомянутом выше понимании).
Важно учитывать, что непропорциональное снижение численности одного из генетических вариантов не только непосредственно изменяет генетическую структуру популяции, но может иметь и далеко идущие последствия.
По этому поводу Б. К. Павлов пишет следующее: «В последнее время [Шварц, 1965] показано, что изменение возрастной структуры ведет к направленному преобразованию генетического состава популяции. В эксперименте удалось обнаружить возрастную избирательность промысла, особенно резко она проявляется среди группы сеголеток. При высокой интенсивности промысла (под промысел попадает 50% популяции) особи с весом сухого хрусталика глаза 25–26 мг (что соответствует рождению в конце марта–начале апреля) остаются в популяции в большем числе. Возрастная избирательность в этом случае вызовет изменение генетической структуры популяции. Эти изменения оказываются еще более существенными в связи с одной особенностью белок этой группы: на следующий год они вступают в размножение ранее, чем белки других возрастов. Среди них количество особей, дающих второй помет, значительно больше, чем в других группах. Некоторые данные позволяют утверждать, что эта особенность связана с наследственными свойствами. Конкретные итоги преобразования популяций будут определяться взаимодействием давления естественного отбора и промысла. Изучение закономерностей преобразования популяции при изменении условий существования и действие промысла открывает пути управления природными популяциями».
Резкие изменения численности автоматически не влекут за собой изменения генетической структуры популяции. Об этом очень ясно свидетельствуют известные наблюдения и опыты Форда [Ford, 1963].
{190}
В нашей лаборатории этой же проблеме было посвящено исследование В. Е. Берегового [1966, 1967]. Изучались семь популяций полиморфного вида пенницы обыкновенной (Philenus spumarius L., Homoptera: Cercopidae). Было установлено, что на Урале встречаются одиннадцать типов окраски, известных под следующими названиями: typica, populi, trilineata, flavicollis, gibha, leucocephala, quadrimaculata, albomaculata, leucophtalma, marginella, lateralis. Все изученные популяции различаются по соотношению частот этих типов окраски, причем различия статистически существенны. Имеющийся материал не позволяет установить прямую связь между различиями мест обитания и особенностями состава популяций. Популяции, обитающие в лесу, так же хорошо отличаются друг от друга, как и популяции леса и луга. Однако наблюдается явная зависимость различий между популяциями от степени изоляции и их взаимного расположения. Последнее связано с миграцией особей между соседними популяциями.
Результаты своих исследований В. Е. Береговой [1967] суммирует следующим образом: «Состав четырех популяций был прослежен с 1964 до 1966 г. Анализ полученных данных показал, что степень различия между популяциями не перекрывается различиями любой из этих популяций от года к году. Каждая популяция сохраняет свои характерные особенности состава на протяжении трех лет. Наблюдаемая стабильность межпопуляционных различий особенно подчеркивает установленный факт резких и несовпадающих колебаний численности, затрудняющих нередко сбор материалов в отдельных популяциях. Эти колебания численности не совпадают даже в соседних популяциях на расстоянии 400 м друг от друга.
Весь имеющийся материал свидетельствует в пользу того, что генофонд каждой популяции формируется в сложной зависимости от факторов среды и проявляет стабильность во времени. Характер межпопуляционных различий на нашем материале обнаруживает большое сходство с различиями, часто наблюдающимися между подвидами».
Эти наблюдения показывают, что популяция обладает способностью поддерживать относительное постоянство своего состава, несмотря на очень резкие колебания численности. Изменение генетического состава популяции, связанное с изменением численности, может быть использовано естественным отбором для быстрых преобразований, соответствующих изменениям среды. В других случаях отбор в течение короткого времени исправляет возникшие нарушения. Популяция не находится во власти слепого случая, определяющего ее генетическую специфику. Изменчивость фенотипической структуры популяций позволяет существовать им в резко различающихся условиях среды.
Все случаи «катастрофической», неизбирательной элиминации, за исключением таких редких, как извержение вулкана и т. п., повторяются во времени, поэтому, приходясь на разные сезоны
{191}
года, оказывают сбалансированное действие на местные популяции. В этом плане и нельзя ожидать, чтобы оседлые популяции обнаружили изменения, подобные описанным. Они уже претерпели подобные изменения столетия и тысячелетия тому назад. Если же популяция попадает в новые климатические условия или (см. ниже) подвергнется воздействию дополнительных сезонно «ориентированных» и повторяющихся из года в год истребительных мероприятий, тогда мы вправе ожидать подобную картину изменения структуры популяции.
Математическое моделирование анализируемой здесь закономерности может быть еще более уточнено. Так, например, может быть учтено повышение скорости полового созревания животных при снижении плотности популяций, увеличение половой активности самцов, изменение смертности перезимовавших животных (производителей) и т. п. Однако подобные уточнения не входят в нашу задачу. Мы стремились показать, что прогресс популяционной экологии создал предпосылки для математического моделирования микроэволюционных процессов, моделирования, значительно более близкого к реальной природной обстановке, чем то, которое легло в основу учения о генетическом дрейфе, генетико-автоматических процессах. При этом становится ясным, что изменение экологической структуры популяций, независимо от того, какими непосредственными причинами оно вызывается (в том числе и неизбирательной элиминацией), имеет следствием изменение ее генетической структуры. Это дает нам право говорить об экологических закономерностях эволюционного процесса.
Мы затронули лишь частный случай проблемы, отметили значение возрастного состава популяций и неизбирательной элиминации, изменяющей внутрипопуляционную структуру. При этом мы сознательно использовали лишь элементарнейший математический аппарат и не стремились описать выявленные закономерности в обобщенных формулах, хотя сделать это было бы не сложнее, чем создать формулы, отражающие скорость преобразования популяций в зависимости от селекционных преимуществ (selektionswerf) отдельных генетических вариантов. Нам важно было показать, что теоретический анализ приводит к выводу о многообразии механизмов направленного преобразования генетической структуры популяций. Индивидуальный отбор, которому до самого последнего времени приписывалась едва ли не монопольная роль в направленном изменении популяций, является лишь одним из таких механизмов.
Теоретический анализ затронутой проблемы должен быть прежде всего использован в качестве программы соответствующих экспериментальных работ (как в лабораторных, так и в полевых условиях), программы, закладывающей основу экспериментальной эволюционной экологии. Результаты этих работ окажут в свою очередь влияние на развитие теоретической экологии. Это сделает современную эволюционную теорию подлинно синтетической.
{192}
Теоретическое значение подобных исследований очевидно. Они имеют не меньшее практическое значение. В настоящее время мы не имеем ни малейшей возможности влиять на ход естественного индивидуального отбора в природе. Но «групповым отбором» мы управлять можем. Зная ход сезонной изменчивости генетической структуры популяции, мы можем относительно просто осуществлять направленное изменение генетической структуры популяции, т. е. фактически управлять микроэволюционным процессом. Более того, мы часто делаем это уже сейчас. Истребительные мероприятия обычно принимают форму избирательной элиминации и всегда приурочены к определенному сезону. По ряду причин особенно эффективны весенние истребительные работы, но при этом изменения генетической структуры популяции неизбежны. Можно полагать, что это приводит к численному преобладанию менее плодовитых, но более жизнестойких особей. Следует подумать: выгодно ли это? Открываются перспективы изменения качества природных популяций. Трудно предвидеть, какие это повлечет последствия для практики борьбы с вредителями и использования полезных животных.
Более того, сама форма проведения истребительных мероприятий приводит к нарушению экологической структуры популяций, что неизбежно ведет и к ее генетическим преобразованиям. При борьбе с грызунами нередко используют препараты мышьяка. Оказалось, что мышьяк оказывает на грызунов избирательное действие. Самки и молодые животные гибнут в относительно меньшем числе, чем взрослые самцы [Junkins, 1963]. Нетрудно представить себе, к каким результатам приведет длительное использование мышьяковистых приманок. В этих условиях селекционное преимущество получают генетические варианты, отличающиеся большой скоростью полового созревания, так как преимущественно за их счет произойдет восстановление популяции. Напрашивается вывод, что применение мышьяка для борьбы с грызунами может в течение короткого времени привести к созданию популяции, отличающейся исключительной скоростью воспроизводства стада.
В данном случае истребительные работы приводят к качественному преобразованию популяции в невыгодную для человека сторону. Если же теоретические исследования достаточно развиты, то будут созданы условия для разработки такой системы истребительных мероприятий, которая не только уменьшит численность вредителей, но и снизит потенциал их воспроизводства. Возможно, что изменение качества популяции окажется более эффективным средством снижения численности вида, чем непосредственные истребительные мероприятия.
Естественно, что это в принципе справедливо не только в отношении истребительных работ, но и промысла. Система промысла определяет не только количественный, но и качественный состав популяции. Приведем несколько примеров.
{193}
Промысел мелких хищников капканами ведет к нарушению нормального соотношения полов в сторону самок. В отдельных случаях эти нарушения могут быть столь значительны, что возникает угроза массового прохолостания самок. В этих условиях явным селекционным преимуществом должны обладать самцы, отличающиеся ранним половым созреванием, так как при недостатке в популяции самцов участие в размножении молодых производителей должно иметь особое значение. Целеустремленные исследования в этом направлении (сравнение скорости полового созревания самцов опромышляемых и неопромышляемых районов) могли бы иметь, таким образом, большое значение и для теории, и для практики. Капканный промысел всегда избирателен, этим и определяется его действие на качественный состав популяции. Иное значение должен иметь промысел, приближающийся к неизбирательной элиминации. Этому условию в большей степени соответствует, например, ружейный промысел белки. При этом чем интенсивнее ведется промысел, тем больше он соответствует неизбирательной элиминации, так как при промысле малой интенсивности в большем числе будут отстреливаться более активные животные, оставляющие больше следов. Поэтому в интенсивно опромышляемых угодьях отбор должен идти на плодовитость. Создается популяция, отличающаяся повышенной плодовитостью.
Приведенные данные показывают, что анализ явлений, объединяемых понятиями «возрастная структура популяций» и «неизбирательная элиминация», может быть использован для развития общей теории эволюции. Не меньшее значение имеет и изучение пространственной структуры популяций.
ПРОСТРАНСТВЕННАЯ СТРУКТУРА ПОПУЛЯЦИЙ — ФАКТОР МИКРОЭВОЛЮЦИИ
Пространственная структура популяций как фактор микроэволюционного процесса уже давно привлекала к себе внимание исследователей. Достаточно вспомнить, что одна из глав синтетической теории эволюции особое внимание уделяет проблеме изоляции (в том числе и неполной) структурных подразделений вида.
В последнее время интерес к изучению пространственной структуры популяций в эволюционном плане в значительной степени возрос в связи с теорией Левонтина [Lewontin, 1965] о популяционном отборе, суть которой заключается в следующем. Если по каким-либо причинам популяции или микропопуляции вымирают, их место занимают другие. При этом новый генный комплекс в новой среде не копирует старый, популяция преображается, происходит эволюционный сдвиг.
Исследование этого процесса наталкивается на ряд трудностей не только технических, но и принципиальных. Дело в том, что, когда говорят о вымирании популяции, имеют в виду не популя-
{194}
цию в строгом смысле слова (экологическое единство, способное самостоятельно регулировать свою численность), а вымирание внутрипопуляционной группы, части единой популяции. Вымирание подобных группировок оказывает на популяцию в целом сложный и противоречивый эффект. Недостаточно строгий подход к понятию «популяция» в данном случае, как и во многих других, может привести к ошибочным представлениям.
Тем не менее исследование явлений, подпадающих под понятие межпопуляционного отбора (interdeme selection), уже в настоящее время содействует лучшему пониманию механизмов эволюционного процесса. В частности, работы этого направления приводят к анализу возможных противоречий между внутрипопуляционным отбором (обычная форма естественного отбора, intrademe selection) и отбором межпопуляционным.
В качестве искусственного примера Левонтин [Lewontin, 1965] указывает на возможность отбора животных, способных к особо эффективному потреблению растительного корма. Подобные животные оставляли бы после себя больше потомства, и вскоре это привело бы к формированию популяции «супер-травоядных» животных. Растительность была бы уничтожена, и популяция вымерла, уступив место другой, использующей наличные запасы корма более экономно.
Свои предположения Левонтин стремится подкрепить и примером из лабораторной и полевой практики [Lewontin, Dunn, 1960; Lewontin, 1962]. В естественных популяциях домовой мыши часто встречается мутация t. В результате внутрипопуляционного отбора частота встречаемости гена t резко увеличивается. Однако самцы, гомозиготные по t, стерильны. В результате производительность популяции падает и она замещается другой. Этот пример Левонтин считает блестящим (excelent) подтверждением своей теории межпопуляционного отбора.
Независимо от Левонтина идея группового отбора нашла выражение в интересной работе А. М. Чельцова-Бебутова [1965], исследовавшего эволюцию тетеревиных токов. Автор считает, что для того чтобы объяснить возникновение брачного ритуала тетерева следует отойти от представлений об индивидуальном отборе и мыслить на уровне группового отбора, на уровне микропопуляций, под которыми он понимает группы птиц, слетающихся на один ток. Усложнение и интенсификация брачного ритуала стимулируют половую активность самок и повышают производительность популяции. «В результате микропопуляции тетеревов с более длительным периодом токования оказывались в более выгодном положении, часть самок могла снести повторные (компенсаторные) кладки, увеличив таким образом общую плодовитость популяции».
Нетрудно заметить, что в разнообразных примерах межпопуляционный отбор выступает как фактор, отсеивающий популяции с неудовлетворительной генетической структурой, а не как творче-
{195}
ский фактор. Это не случайность, определяющаяся неудачным подбором примеров, а закономерность. Говорить о межпопуляционном отборе можно лишь условно, так как значение вымирания (полным оно никогда не бывает) локальных «популяций» может быть оценено только на основе исследования процессов, происходящих в популяции в целом, популяции в строгом понимании этого слова. Правильнее говорить не о межпопуляционном отборе, а о роли динамики пространственной структуры популяции в микроэволюции. При этом под пространственной структурой популяции мы понимаем особенности распределения животных по территории, в том числе и возникновение локальных микропопуляций, заселяющих своеобразные по условиям среды биотопы.
Анализ рассматриваемого вопроса должен основываться на двух основных фактах. Первый из них: колебания численности вида захватывают громадные территории. Примеры в данном случае совершенно излишни, так как массовое размножение многих видов, относящихся к различным таксономическим группам, многократно описывалось и всесторонне исследовалось. Пик численности (или депрессия) распространяется нередко не только на целые популяции, но и на группы популяций. Во время вспышки численности происходит заселение территорий, в которых вид или отсутствует, или встречается в обычные годы в незначительном числе. При этом неизбежно происходит перемещение популяции или резкое увеличение той территории, в пределах которой фактически осуществляется панмиксия.
Столь же обширна территория, в пределах которой проявляется депрессия численности вида. Нередко она охватывает целые ландшафтные зоны. Так, падение численности лемминга или песца в отдельные годы наблюдается по всей необъятной территории Полярной Евразии. Во время депрессии численности животные сохраняются лишь в стациях переживания (в понимании Н. П. Наумова [1963]) или на отдельных участках ареала вида, где по каким-либо причинам складываются более благоприятные условия существования. Все сказанное — азбучные истины современной популяционной экологии животных, по эти истины еще не используются в полной мере для анализа экологических механизмов микроэволюционного процесса.
Другая группа фактов, имеющих особое значение для оценки возможной роли межпопуляционного отбора, уже не может быть отнесена к азбуке экологии. Речь идет о процессе, противоположном только что описанному. Если пик численности и резкая ее депрессия охватывают значительные территории, то в обычные («средние») годы колебания численности даже соседних микропопуляций могут быть резко различными. Популяции полевок в лесных колках, кустарниковых зарослях, на залежах, лугах, полях подвергаются различным воздействиям среды даже в том случае, если они расположены в непосредственной близости, в одной и той же географической среде. В силу этого динамика их числен-
{196}
ности неизбежно должна быть различной. То, что это действительно так, показывают многочисленные исследования, но особенно ясно — работа Хайна и Томсона [Hayne, Thomson, 1965]. Они в течение 10 лет изучали динамику численности Microtus pennsylvanicus в штатах Мичиган и Висконсин. Наблюдения проводились на 197 стационарных пунктах. Среднее расстояние между пунктами — 10 км. Математическая обработка материала показала, что корреляция между изменениями численности в соседних популяциях или статистически недостоверна, или хотя и реальна, но столь слаба (r=0,28), что не может быть принята во внимание при проведении истребительных работ. Делается обоснованный вывод: в пределах одного района колебания численности вида подчиняются сходным закономерностям, но локальная численность (численность отдельных популяций или микропопуляций) может резко отличаться от средней по району и от численности соседних поселений.
В нашей лаборатории аналогичные исследования были проведены на ондатре В. Ф. Сосиным [1967]. В течение трех лет изучалась динамика численности ондатр на нескольких озерах, расположенных на расстоянии 6–10 км друг от друга в лесостепи Зауралья. Установлено, что колебания численности ондатр, даже на водоемах, сходных по своему характеру, несинхронны. В то время как на одном из них количество животных возросло примерно с 250 до 450, на другом имело место ее падение с 350 до 140 ондатр. На третьем озере численность во все годы наблюдения изменялась в значительно меньших пределах, примерно от 120 до 160 ондатр. Движение численности сопровождалось изменениями возрастной структуры. В поселениях, где количество ондатр по сравнению с предшествующим годом увеличилось, наблюдается рост числа молодых, приходящихся на каждую размножавшуюся самку (с 5,4 до 9,4 на одном озере, с 6 до 20 — на другом). На водоеме же, где численность в разные годы менялась слабо, возрастной состав варьировал в пределах 12–16 сеголеток на одну размножавшуюся самку. Сопоставление числа производителей с интенсивностью размножения и плотностью осенней популяции приводит к выводу о том, что в рассматриваемом случае динамика численности определялась в основном выживаемостью молодняка (миграций в период наблюдений не отмечалось).
Приведенные данные показывают, что поселения ондатр на озерах, расположенных в непосредственной близости, подчиняются различным закономерностям динамики численности и могут рассматриваться как микропопуляции с характерными особенностями движения населения и возрастной структуры.
Мелкие мышевидные грызуны превосходно плавают и могут переплывать даже довольно широкие и быстрые реки. Тем не менее несомненно, что обмен особями между популяциями полевок (за исключением, естественно, видов, ведущих полуводный образ жизни) ограничен и осуществляется, вероятно, Далеко не каждый
{197}
год. В этом отношении интересен такой пример. Нами была обследована популяция экономок (М. oeconomus) на берегу большого степного озера. На озере много сплавинных островов, которые в летнее время представляют собой защищенные от врагов и богатые кормом места обитания полевок. Острова расположены на расстоянии всего около 100 м от берега, а расстояние между отдельными островами еще меньше. В зимнее время условия существования на островах хуже, чем на «материке», так как почва промерзает. Зимой большинство островных популяций вымирает. Это дает возможность составить представление о степени изоляции полевок, разделенных водной преградой порядка десятков и сотен метров. Оказалось, что, хотя экономка могла бы достичь любого острова за десяток минут, острова заселяются далеко не каждый год. Были поставлены и специальные эксперименты. Маркированные экономки завозились на острова и выпускались. Оказалось, что преобладающее большинство из них остается в местах выпуска и не переплывают с острова на остров или на берег.
Аналогичные по существу наблюдения у В. Ф. Сосина [1967]. Проведенная им маркировка показала, что ондатры не часто переходят с озера на озеро даже в том случае, если расстояние между ними измеряется десятками метров. Много подобных примеров можно было бы привести из литературы. Они показывают, что единая, казалось бы, популяция распадается на группы частично изолированных популяций, обмен генами между которыми в значительной степени ограничен.
Вполне аналогичные данные могут быть приведены и по другим видам. Для иллюстрации воспользуемся данными заготовок ряда видов пушных зверей на Ямале и в прилегающих районах Урала. Из богатого материала, представленного Г. Е. Рахманиным [1959], приведем лишь несколько примеров. В 1954 г. в целом по округу численность ондатры по сравнению с предшествующим годом почти не возросла (164813 заготовленных шкурок против 155902), но в Ямальском районе она возросла в 3 раза, в Надымском — более чем вдвое, а в Красноселькупском снизилась. Численность белки в 1956–1957 гг. в Надымском районе снизилась в 1,5 раза, а в Ямальском увеличилась в 20 (!) раз. В эти же годы численность горностая в Красноселькупском районе увеличилась в 2 раза, а в Шурышкарском снизилась втрое. В предшествующие годы происходило обратное: в Шурышкарском районе численность несколько увеличилась, в Красноселькупском снизилась более чем в 4 раза. Столь же несинхронными были колебания численности в разных районах Ямала и других видов.
Несмотря на то что данные заготовок нельзя считать вполне строгим показателем движения численности, они совершенно неоспоримо показывают, что соседние поселения животных нередко характеризуются разными закономерностями динамики численности.
{198}
Изменение численности имеет следствием неизбежное изменение генетического состава популяции. Это изменение может быть существенным или ничтожно малым, но оно не может не произойти. Об этом свидетельствуют твердо установленные законы популяционной генетики. Отсюда следует, что конкретный эффект обмена генами между популяциями в значительной степени определяется сравнительным ходом динамики численности животных. О значении этого явления мы будем подробнее говорить ниже.
Другая сторона той же проблемы — вымирание микропопуляций. Интенсивность размножения вида соответствует условиям его существования. Чем выше смертность — тем выше рождаемость. Не случайно интенсивность размножения слонов в тысячи раз слабее, чем интенсивность размножения мышей! Из этой, опять-таки азбучной истины экологии следует, что для того чтобы популяция вымерла, совсем не обязательно катастрофическое сочетание внешних факторов. Если по каким-то причинам смертность начинает превышать рождаемость, то вымирание популяции — это лишь вопрос времени. Когда речь идет о животных-эфемерах — это вопрос очень незначительного промежутка времени. Тем не менее в большинстве подобных случаев вымирания не происходит, так как при резком сокращении численности популяции увеличивается число иммигрантов из соседних популяций. Однако когда популяция замкнута, то легко обнаруживается, что незначительное снижение нормального темпа воспроизводства популяции обрекает ее на гибель. Это было ясно показано на островных популяциях домовой мыши [Lidicker, 1966]. Оказалось, что при совместном обитании с Microtus californicus нормальная беременность мышей нередко нарушается. Это ведет к снижению темпов воспроизводства популяции и ее быстрейшему и полному вымиранию, несмотря на «обычную» смертность и в целом высокую интенсивность размножения.
Так как подобное изменение темпа размножения несомненно встречается в природе весьма часто, то отсюда следует что иммиграция особей из одной популяции в другую — явление неизмеримо более частое, чем это фиксируется прямыми наблюдениями. Однако интенсивность обмена особями подчиняется достаточно строгим закономерностям, о которых мы только в последнее время начинаем получать точную информацию. С другой стороны, все чаще поступают данные, показывающие, что даже соседние популяции — это довольно замкнутые экологические системы. До тех пор, пока популяция полностью сохраняет свою жизнеспособность, она активно ограничивает проникновение особей извне. Известны и конкретные экологические механизмы, препятствующие обмену особями между популяциями и внутрипопуляционными группами животных. Эти механизмы имеют в своей основе сложную внутрипопуляционную систему господства — подчинения. В этой системе «чужак» лишь в редких случаях может войти в группу особей-доминантов и конкурировать с хозяевами в интенсивном размно-
{199}
жении. Возможно, что имеют определенное значение и иные механизмы изоляции: своеобразие подбора пар (подробнее см. ниже), нарушение беременности у самок при внезапном появлении чужих самцов [Bronson, Eleftherion, 1963], инбридинг [Scossiroli, 1962], стремление животных избегать не освоенных видом территорий [Haggerty, 1966]. Возможно, что у высших животных играет роль и привязанность к колонии, и привязанность отдельных особей друг к другу ([Penney, 1964], наблюдения на пингвинах). Исследование подобных механизмов генетической изоляции только начинается, однако в их существовании нельзя сомневаться. Трудно подобрать лучший пример, чем исследования польских экологов, изучавших микропопуляции мышей на чердаке и в подвале двухэтажного дома [Adamczyk, Petrusewicz, 1966]. Было установлено, что разный тип динамики численности надежно изолирует эти группы животных, что в конечном итоге приводит к возникновению между ними генетических различий (результаты опытов по трансплантационному иммунитету). Можно было бы привести довольно много аналогичных примеров, указывающих на относительную изоляцию группировок животных. Приток особей из соседних популяций резко увеличивается в периоды локальных депрессий численности. По понятным причинам в этих условиях роль иммигрантов в преобразовании генетической структуры популяции существенно возрастает, так как их относительное обилие в новой, формирующейся популяции резко увеличивается. Более того, оказалось, что смертность иммигрантов обратно пропорциональна численности популяции [Andrzeijewski et al., 1963]. Смешение популяций во время депрессии численности имеет поэтому особенно важное значение. Результаты опытов Петрусевича подтверждаются исследованиями Андерсона [Anderson, 1966], показавшего, что между несколькими соседними поселениями мышей длительное время не происходило обмена генетическим материалом.
Сказанным не исчерпываются те важнейшие экологические закономерности, знание которых необходимо для правильной оценки роли пространственной структуры популяции в микроэволюции. Большая серия работ, на которые мы имели возможность сослаться в главе V, ясно показывает, что перемещение животных разного пола и возраста различно. Новые факты в этом отношении были получены Петрусевичем с сотрудниками [обзор данных см. Petruseewicz, Andrzeijewski, 1962], которые считают даже возможным выделить «мигрантов» и «оседлых» в экологически различные группы особей.
Падение численности животных имеет различные причины, но в относительно очень большом числе случаев при резком снижении численности популяции в первую очередь сокращается число рекрутируемых в популяцию молодых (следствие падения интенсивности размножения при увеличении детской смертности). Это значит, что при замещении одной популяции другой происходит
{200}
преимущественное смешение старых особей одной популяции с молодыми пришельцами. Важная задача эволюционной экологии заключается в том, чтобы установить конкретные проявления этой закономерности, но в ее если не всеобщем, то во всяком случае широком распространении вряд ли можно сомневаться.
После этих предварительных замечаний перейдем к анализу роли динамики пространственной структуры популяции в преобразовании ее структуры генетической. Как уже упоминалось, это влияние противоречиво. Логический анализ показывает, что сложность пространственной структуры популяций имеет следствием различный ход динамики численности отдельных микропопуляций, способствует сохранению единства популяции и стабилизации ее генетического состава (при условии постоянства направления и интенсивности естественного отбора). Этот вывод кажется парадоксальным, но он непосредственно вытекает из известных положений (см. гл. II) о связи между динамикой численности и изменением генетического состава популяций. Резкое снижение численности может привести к потере определенного гена в пределах отдельных микропопуляций, но допустить, что в двух соседних популяциях характер обеднения генофонда случайно окажется одинаковым, совершенно невозможно. Поэтому перемешивание микропопуляций и популяций неизбежно приводит к восстановлению исходного генофонда, и под влиянием отбора исходное соотношение разных генетических вариантов будет восстановлено в течение нескольких поколений. Об этом свидетельствуют приведенные нами в главе II экспериментальные данные.
В качестве конкретного примера мы можем воспользоваться уже описанными нами исследованиями динамического полиморфизма остромордой лягушки. В Курганской области в 1966 г. исследовались две микропопуляции. Генетический состав животных младшего возраста в пространственно изолированных популяциях оказался различным: процент striata в одной из них оказался равным 43,9±3,7, в другой — 55,3±3,46. Однако лягушки старшего возраста оказались неразличимыми (процент striata 66,7±7,4 и 66,0±6,9). Изменение генетического состава отдельных микропопуляций, вызванное неизвестными нам изменениями среды, не привело к их дифференциации, так как в процессе миграции взрослых животных произошла консолидация популяции в строгом смысле этого слова в единое генетическое целое.
Таким образом, сложная пространственная структура популяций не влечет за собой неминуемую генетическую или морфофизиологическую дифференциацию вида.
Очень многие мелкие виды ландшафтных зон образуют бесчисленное число популяций, но это не ведет к интенсивному внутривидовому формообразованию. Еще раз вспомним сибирского лемминга. Его дифференциация выражена столь слабо, что Сидорович [Sidorowicz, 1960] счел возможным объединить всех материковых леммингов Азии в один подвид. Ему возражают
{201}
В. Г. Кривошеев и О. Л. Россолимо [1966], но выделяют в Евразии два подвида, из которых один (Lemmus sibiricus chrisogaster) более чем сомнителен.
Область распространения красной полевки (Clethrionomys rutilus) громадна. На северном и южном пределах ареала вид представлен полуизолированными популяциями, приуроченными к островкам леса среди степной или тундровой растительности. Тем не менее, как показали тщательные исследования В. Н. Большакова [1962], заслуживают таксономического закрепления всего лишь 4 подвида, отличающиеся преимущественно по окраске [Большаков, Шварц, 1962]. Этот пример особенно показателен, так как ареал красной полевки охватывает ряд ландшафтных зон — от степи до тундры.
Для многих видов мышевидных грызунов описано множество подвидов, но большинство из них выделяется на основе ничтожных (нередко мнимых) отличий по окраске или пропорциям тела и черепа. Не случайно каждая ревизия внутривидовой систематики мышей или полевок многократно сокращает число подвидов. Между тем подвиды многих крупных млекопитающих — действительно резко дифференцированные формы. Достаточно сравнить структуру вида той же красной полевки со структурой вида большинства копытных [новейший обзор см. Гептнер и др., 1961], чтобы различие в формообразовании крупных и мелких млекопитающих стало очевидным.
Это различие трудно объяснить на основе ставших традиционными представлений синтетической теории эволюции, так как мелкие виды образуют неизмеримо большее число относительно изолированных популяций, чем крупные. Между тем если к внутривидовой таксономии грызунов подойти с тем же масштабом, что и к таксономии копытных, то многие их виды с громадным ареалом (ондатра, полевка-экономка, водяная полевка, красная полевка и мн. др.) должны были бы быть признаны мономорфными, а лось, косуля, благородный олень и другие виды копытных образуют подвиды, вокруг которых все еще не прекращаются споры относительно их возможной видовой самостоятельности.
Отсюда следует, что скорость внутривидовой дифференциации определяется в первую очередь условиями существования, в процессе приспособления к которым и происходит дивергенция разных форм вида. Для мелких млекопитающих, в жизни которых ведущее значение имеет микроклимат, условия существования в лесотундре и лесостепи могут оказаться весьма сходными, для лося же они резко различны. Поэтому подвиды полевок дифференцированы слабее, чем подвиды лося, несмотря на то что генетико-автоматические процессы должны были бы привести к диаметрально противоположным результатам.
При изменении условий существования мелких млекопитающих изменяются и их морфологические особенности, даже в том случае, если сравниваемые популяции расположены в непосред-
{202}
ственной близости и изоляция между ними слабая. Хороший пример — так называемая биотопическая изменчивость, суть которой заключается в том, что нередко животные (большинство исследований проводилось на грызунах) из разных биотопов одного географического района отличаются между собой значительнее, чем животные разных подвидов [Поляков и др., 1958, и др.]. Сам факт биотопической изменчивости несомненен, но к ее анализу следует подходить с осторожностью.
В большом числе случаев различия между животными из разных биотопов объясняются фенотипической изменчивостью. Приведенные в главе III материалы показывают, что изменение системы коррелятивных связей в развитии индивидов может привести к изменению под непосредственным влиянием среды даже, казалось бы, наиболее стабильных признаков. В отдельных случаях можно допустить, что биотопическая изменчивость имеет в своей основе отличия в генетической структуре микропопуляций. Эти отличия могут быть результатом преобразования популяций в течение одного поколения, а отнюдь не результатом их самостоятельной эволюции. То, что результаты отбора в одном поколении могут привести к существенным изменениям, доказывается как специальными исследованиями, так и в особенности многочисленными наблюдениями, относящимися к проблеме «сезонного отбора». Таким образом, и «биотопическая изменчивость» не противоречит утверждению об отсутствии неизбежности внутривидовой дифференциации при усложнении пространственной структуры популяций животных.
Сложная топографическая структура популяции лишь в особых случаях приводит к дифференциации популяции и к последующему формообразованию, но нам представляется, что по крайней мере во многих случаях она является мощным фактором эволюции популяции как единого целого. Сложная топографическая структура популяции выступает не только как стабилизирующий и объединяющий, но и как творческий фактор. К этому выводу нас приводит синтез изложенных выше данных по экологии популяций с данными по популяционной генетике.
Один из важнейших выводов популяционной генетики сводится к тому, что большинство межпопуляционных отличий — полигенной природы. При этом один и тот же признак может иметь различную генетическую основу, а совместное действие разных генов оказывает взаимно усиливающий эффект (адаптивное действие генов).
По первому вопросу наиболее важный материал дают уже упоминавшиеся нами исследования по воздействию ядов на насекомых. Показано (сводку данных см. [Crow, 1957; Milani, 1957; Brown, 1958; Benett, 1960, и др.]), что у разных видов насекомых генетические механизмы возникновения ядостойких популяций различны. Более того, в разных лабораториях возникновение ядостойких «рас» имеет различную генетическую основу. У некоторых
{203}
линий дрозофилы резистентность оказалась связанной с одним доминантным геном [Ogaki, Tsukamoto, 1957], у других — с адаптивным действием многих генов [Crow, 1957]. Беннет [Benett, 1960] указывает, что во многих случаях эти различия между штаммами дрозофилы определяются различным характером воздействия ДДТ на экспериментальные популяции (аэрозоли, ДДТ на бумаге, воздействие яда, примешанного к пище личинок и т.п.). Однако крайне важно, что и в одинаковых условиях у линий одной лабораторной колонии резистентность к ядам нередко возникает на различной генетической основе.
Эти наблюдения и эксперименты делают очевидным, что и в природных условиях приспособления к одинаковым условиям в пределах разных микропопуляций могут происходить и несомненно происходят на разной генетической основе. Об этом свидетельствуют как прямые наблюдения, на основе которых возникло представление о том, что тождественные фенотипы могут определяться разными генотипами [Espinasse, 1964], так и многие косвенные указания. Особенно интересны экспериментальные работы, проведенные на птицах, показывающие, что строгого соответствия между фенотипической и генотипической близостью животных нет [Hall, 1963]. Этот вывод приобретает особое значение в свете общих закономерностей аддитивного действия генов. Гены аддитивного действия работают как единая система, но каждый из них усиливает выражение определенного признака, и его эффект не зависит от присутствия или отсутствия других генов [Dobzhansky, 1955].
Очень важно, что адаптивный характер генетических различий обнаруживается и по таким биологически существенным признакам, как скорость роста и вес тела [Кирпичников, 1967].
На Всесоюзном совещании по отдаленной гибридизации растений и животных (Москва, 1968 г.) особо отмечалось, что большинство различий между скрещиваемыми формами наследуется аддитивно [Кузьминых, 1968].
Аддитивный эффект генов — это лишь один из частных случаев проявления их совместного действия, но в рассматриваемом здесь плане он представляет особый интерес. Дело не только в том, что на основе описанных экологических механизмов аддитивное действие генов может привести к резкому увеличению темпов эволюционных преобразований. Оно может привести к совпадению направления отбора и изменчивости. Этот вопрос занимает умы исследователей еще со времен Дарвина, но, как нам кажется, не получил еще удовлетворительного решения. Если допустить, что в результате отбора определенного давления в соседних микропопуляциях получат распространение фенотипически сходные, но генетически различные варианты, то при объединении микропопуляций возникают условия для совмещения в едином генотипе генов, обладающих сходным фенотипическим проявлением. Если действие этих генов суммируется, то в результате объединения
{204}
микропопуляций в единую популяцию возникнут новые генотипы, обладающие более сильным фенотипическим выражением даже в том случае, если давление отбора снизится или прекратится. Значение подобных явлений трудно переоценить, но в настоящее время развиваемые взгляды могут быть обоснованы преимущественно теоретически. Их конкретизация на основе полевых и лабораторных исследований представляется нам важной задачей эволюционной экологии. Как указывалось, далеко не всегда объединение двух генов сходного фенотипического проявления в едином генотипе приводит к усиливающему эффекту. Вероятно, значительно чаще их совместное действие приводит к появлению новых свойств организмов. В таких случаях смешение микропопуляций приведет к иным, но не менее важным следствиям. Произойдет обогащение общего генофонда популяции, ее возможности более полного и быстрого приспособления к среде увеличиваются.
Этот важный вывод может быть в настоящее время подкреплен экспериментальными данными. Левонтин и Берч [Lewontin, Birch, 1966] изучили процесс преобразования модельных популяций австралийских плодовых мух рода Dacus. Предварительные наблюдения в природе показали [Birch, 1961], что за последнее столетие D. tryoni расширили свой ареал благодаря приспособлению к высоким температурам. Было установлено, что расширению ареала предшествовала гибридизация D. tryoni с очень близким видом («почти видом», near-species) D. neohumoralis (нарушение репродуктивной изоляции произошло в результате обогащения кормовой базы обоих видов новыми видами фруктов). Авторы предположили, что быстрое приспособление D. tryoni к новым условиям существования связано с обогащением генофонда. Эта гипотеза была проверена экспериментально. В течение двух лет чистые культуры D. tryoni и гибриды D. tryoni X D. neohumoralis содержались при температуре 20, 25 и 31,5° С. В течение первого года чистая культура оказалась продуктивнее гибридной, но к концу второго года при температуре 31,5° гибриды оказались более жизнеспособными и продуктивными чем, D. tryoni. Авторы справедливо заключают, что результаты опытов свидетельствуют о том, что внедрение в популяцию генов, которые сами по себе не адаптивны, ведет к быстрой адаптивной эволюции. Это позволяет думать, что временное разъединение популяций (или микропопуляций) вида, во время которого под влиянием экологических механизмов происходит их генетическая дифференциация и их последующее воссоединение, является мощным фактором эволюции. И это важное положение может быть подтверждено прямыми экспериментами, проведенными в нашей лаборатории группой исследователей. А. В. Покровский длительное время изучал экологические и морфофизиологические особенности двух подвидов узкочерепной полевки и их гибридов. Н. А. Овчинникова проделала аналогичную работу с полевкой-экономкой. Оба вида были детально изучены в природе О. А. Пястоловой, К. И. Копейным,
{205}
В. Н. Бойковым и автором. Результаты этих работ опубликованы в серии статей [Копеин, 1958; Шварц и др., 1960; Овчинникова, 1965; Покровский, 1967]. Были установлены закономерности наследования отдельных признаков и показано, что важнейшие морфологические и экологические особенности форм вида наследственны. В. Г. Ищенко [1967] обработал полученные данные с помощью аллометрического метода и показал, что при скрещивании двух подвидов гибриды нередко обладают новыми признаками, и, что особенно важно, эти «новые признаки» приводят к дифференциации вида.
<![if !vml]>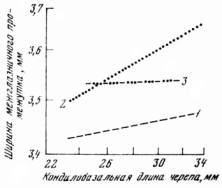 <![endif]>
<![endif]>
Рис. 15. Относительный рост ширины межглазничного промежутка у южного (Microtus о. oeconomus) (1), северного (М. о. chahlovi) (2) подвидов полевки-экономки и их гибридов (3)
Одно из наиболее существенных отличий северной формы экономки (М. о. chahlovi) от южной — разный соотносительный рост ширины межглазничпого промежутка (по отношению к длине черепа). Аллометрический экспонент южного подвида 0,040±0,006, северного 0,118±0,006. Другими словами, у северного подвида при изменении длины черепа межглазничная ширина увеличивается быстрее, чем у южного. У гибридов первого поколения увеличение длины черепа практически не приводит к увеличению его ширины (рис. 15). Еще более интересный материал дает изучение наследования длины черепа. Аллометрический экспонент обоих подвидов одинаков (соответственно 0,461+0,006 и 0,468+0,003), у гибридов—существенно различен (0,405±0,012). Аналогичный результат дали опыты по гибридизации подвидов узкочерепной полевки. Зависимость веса почек от веса тела у северного (М. g. major) и южного (М. g. gregalis) подвидов одинакова (соответственно 0,535 и 0,511). У гибридов аллометрический экспонент 0,779 (!). Длина черепа северного подвида по мере роста животных нарастает быстрее, чем у южного (соответственно 0,431 и 0,394; у гибридов — 0,501). Этот пример особенно интересен. Можно считать доказанным, что северный подвид произошел от южного [Шварц, 1961]. В числе его отличий — своеобразная скорость рос-
{206}
та черепа. Однако гибридизация приводит к еще более резкому увеличению скорости его роста.
Принципиально сходные наблюдения были проведены на трясогузке Motacilla flava. Оказалось, что гибридные самцы М. f. flava X M. f. thunbergi отличаются повышенной холодостойкостью [Sammalisto, 1968].
С точки зрения генетики приведенные факты ничем не примечательны, так как известно, что гибриды нередко обладают новыми свойствами по сравнению с исходными формами (см., например, [Дубинин, Глембоцкий, 1967]). Известен и генетический механизм, лежащий в основе этого явления (гетерозиготность по многим генам). Однако в эволюционном плане приведенные примеры заслуживают внимания. Они показывают, что генетические различия между формами одного вида таковы, что при скрещивании возникают генотипы, отличающиеся более сильным выражением отдельных признаков. Еще важнее, что «новые» признаки обнаруживаются и при скрещивании внешне одинаковых форм. И в данном случае генетическая природа явления ясна: фенотипически одинаковые формы могут развиваться на основе разных генотипов. Это показывавает, однако, что перемешивание разных популяций может привести к резким морфофизиологическим сдвигам. Само собой понятно, что конечный эволюционный результат этих сдвигов определяется отбором в гибридной популяции на фоне расщепления признаков. Тем не менее вряд ли можно сомневаться в том, что если не во всех, то во многих случаях разъединение и последующее воссоединение популяций являются мощным фактором акцелерации начальных стадий микроэволюции. Для характеристик и тех процессов, с которыми сталкивается зоотехник при скрещивании пород домашних животных, воспользуемся цитатой из книги Н. П. Дубинина и Я. Л. Глембоцкого [1967]: «Так как породы имеют сложное и полигетерозиготное строение генотипа, то при межпородных скрещиваниях возникает очень сложное полигибридное расщепление, причем большинство выщепляющихся генотипов носит более или менее промежуточный характер. На этом явлении строится воспроизводительное скрещивание. Именно этим обосновал профессор П. Иванов [1935] свои работы по выведению новых пород овец и свиней. Он писал: «...чем большим количеством генов обусловливается какой-либо признак, тем более тонким, т. е. менее заметным становится расщепление в F2 и тем реже появляются вследствие расщепления животные, близкие к исходным формам. Генетик Ланг вычислил, что если, например, какой-либо признак обусловливается действием пяти генов, то на 1024 животных в F2 придется лишь по одному животному, представляющему расщепление в исходные формы, и 912 животных будут с более или менее промежуточным характером. При расщеплении по 10 парам генов, не имеющих полностью доминантного характера, число различных генотипов будет 59049, а на 1043776 особей будет получено всего две особи, го-
{207}
мозиготные по всем 10 парам доминантных или рецессивных генов. Нужно, однако, отметить, что относительно промежуточные по генотипу особи в силу явлений доминирования и взаимодействия генов далеко не всегда будут иметь и промежуточный фенотип. Очень часто при этом могут получаться и фенотипы совершенно новые. В этом одновременно и главная трудность и большие творческие возможности, скрытые в этом методе».
Можно возразить, что в примере, заимствованном мною у Левонтина и Берча [Lewontin, Birch, 1966], гибридизация имела место между «почти-видами», а в наших примерах—между резко выраженными подвидами, прошедшими длительный путь самостоятельного развития. Достаточно ли отличий, которыми характеризуются соседние популяции или микропопуляции? Попытаемся ответить на этот вопрос на основе накопленного в пашей лаборатории материала. И в данном случае воспользуемся показателями аллометрического роста, поскольку они в значительно большей степени характеризуют генетические отличия между популяциями, чем средние величины [Frick, 1961; Ищенко, 1967].
<![if !vml]> <![endif]>
<![endif]>
Рис. 16. Относительный рост сердца полярных крачек популяций Северного (1) и Южного (2) Ямала
Изучение экономки (см. ранее) и двух видов крачек (рис. 16) показало, что и между соседними популяциями, смешение которых вполне реально, по ряду показателей существуют столь существенные различия, что есть все основания думать, что их гибридизация привела бы к столь же резкому эффекту, что и скрещивание подвидов. Обратим особое внимание на рис. 16. Oн показывает, что между популяциями крачек с северного и южного Ямала наблюдаются принципиальные отличия в характере роста важнейшего органа — сердца, которые сигнализируют о различиях во взаимоотношениях этих популяций со средой [Ищенко, Добринский, 1965; Добринский, 1966]. Эти популяции разделены значительным пространством, но изменение условий существования (например, потепление климата) может привести к ослаблению гнездового консерватизма и перемешиванию этих популяций, что привело бы к тем же последствиям, что и гибридизация подвидов.
{208}
Мне кажется, что мы имеем основание говорить о недостаточно оцененном факторе эволюции: динамике структуры популяций. Динамика структуры популяций как фактор эволюции весьма сложна. Она слагается по крайней мере из следующих элементов:
а) изменение возрастной структуры популяций приводит к изменению их генетической структуры, которое может быть усилено элиминацией, кажущейся неизбирательной (экологические механизмы преобразования популяций);
б) динамика пространственной структуры популяций приводит к временной изоляции микропопуляций. Период их временной изоляции может оказаться достаточным для возникновения между ними генетических различий. При воссоединении и перемешивании популяций под воздействием естественного отбора возникают новые сбалансированные генетические системы. В определенных условиях это приводит к быстрому прогрессивному развитию популяции, к резкой акцелерации микроэволюции.
В связи с изучением роли пространственной структуры популяций в преобразовании их генетического состава нужно затронуть вопрос о влиянии хищника на эволюцию жертвы. Всесторонний анализ этой обширной проблемы не входит в нашу задачу, тем более что в общем плане она достаточно полно исследована. «Приспособления хищника влекут за собой контрприспособления жертвы» — это положение стало хрестоматийным. Тем не менее хищник может играть в преобразовании популяции жертвы специальную роль, которая, как нам кажется, осталась незамеченной до настоящего времени.
Когда оценивают влияние хищника на численность его жертвы, сопоставляют число уничтожаемых животных с их общим количеством в обследуемом районе. При этом обнаруживается, что, за крайне редкими исключениями, хищник снимает лишь ничтожную часть «урожая» жертвы. Этот вывод особенно хорошо обоснован наблюдениями над хищными птицами. Обобщение многочисленных данных, проведенное недавно В. М. Галушиным [1966], показало, что «величина изъятия» хищными птицами дичи измеряется единицами или даже долями процентов их поголовья. Возможно, что хищники-миофаги поедают относительно большее число грызунов, но хорошо известно, что даже исключительное обилие хищников не может существенно повлиять на изменение численности процветающих популяций мышей или полевок. То же самое можно сказать и в отношении насекомоядных птиц и насекомых.
Иная картина вырисовывается в том случае, когда влияние хищника оценивается исходя не из общей численности жертвы, а из ее численности в охотничьих участках доминирующих видов. Тот же В. М. Галушин [1966] показал, что величина охотничьих участков хищных птиц находится в прямой зависимости от обилия пищи и колеблется в соответствии с этим в 50–100 раз. Это свидетельствует о том, что большого резерва кормов в пределах охотничьих участков нет; хищник резко снижает численность
{209}
жертвы на отдельных участках даже в тех случаях, когда на ее общую численность он не оказывает никакого влияния. Подтверждением этого могут служить превосходные по точности наблюдения М, Д. Зверева [1930], который установил, что в пределах своих охотничьих участков пустельга уничтожает полевок практически полностью (результаты опытов с установкой шеста для хищных птиц). В отношении насекомых подобные наблюдения нам не известны, но некоторые косвенные данные делают вероятным, что и насекомоядные птицы (подобно хищным) значительно снижают численность своих жертв в пределах гнездовых участков. Об этом свидетельствует смена охотничьих участков отдельными семьями немедленно после того, как птенцы способны покинуть гнездо. Подобные наблюдения были проведены сотрудниками нашей лаборатории на птицах тундры. Так, например, луговые и краснозобые коньки переводят нелетных еще птенцов на новые участки, где продолжают их выкармливать.
Можно считать установленным, что хищник создает своеобразный вакуум в пределах освоенной его жертвой территории. В процессе заселения подобных участков с резко пониженной численностью наблюдаются закономерности, которые имеют непосредственное отношение к обсуждаемой здесь проблеме. Склонность и способность к миграциям у разных внутрипопуляционных групп различна. Если для примера воспользоваться грызунами, то можно быть уверенным, что заселение освободившихся территорий будет происходить в таком порядке: молодые самцы, молодые самки, взрослые самцы, взрослые самки. Субдоминанты будут оседать в «экологическом вакууме» в большем числе, чем животные, занимающие в популяции доминирующее положение.
Таким образом, в результате деятельности хищников даже в идеальной гомогенной популяции возникают микропопуляции с измененной возрастно-половой структурой. Если принять нашу исходную посылку, согласно которой экологически различные группы животных различны и генетически, то отсюда следует, что к концу сезона размножения подобные микропопуляции будут различаться своей генетической структурой и при их воссоединениях с соседними микропопуляциями будут иметь место все те явления, которые сопутствуют смешению генетически различных групп животных.
Сопоставление выводов популяционной генетики с наблюдениями, характеризующими динамику топографической структуры популяций, приводит к некоторым существенным заключениям, касающимся общих механизмов преобразования популяции.
Один из возможных вариантов: несмотря на различия в конкретных условиях существования в разных биотопах, разные микропопуляции подвергаются сходным силам отбора в соответствии с его генеральным направлением в данной географической среде. Условия, в которых живут тундровые грызуны по берегам рек, по склонам холмов, на равнинной тундре, в зарослях кустар-
{210}
ников,— различны, но все они должны обладать приспособлениями к ведущему фактору макросреды — короткому и холодному лету. Это же самое справедливо в отношении любых ландшафтных зон и их хорологических подразделений. Условия существования зеленой жабы в степных водоемах разных типов (хороший пример микропопуляций) различны, но любые популяции этого вида в засушливом климате должны обладать высокой способностью к миграциям, вызываемым частым пересыханием водоемов. Популяции зеленой жабы не могли бы существовать в степи и тем более в полупустыне, если бы они не обладали способностью использовать для размножения временные водоемы. Подобных примеров можно было бы привести очень много.
Различное конкретное проявление динамики численности разных микропопуляций при общем направлении отбора для популяции в целом должны привести к тому, что при восстановлении временно изолированных микропопуляций или популяций вида одного географического района их «достижения» суммируются, усиливаются и делаются достоянием популяции в целом. В других случаях «приобретения» микропопуляции не суммируются, но создают основу для интеграции новых генетических вариантов, более полно приспособленных к условиям географической среды. В этом процессе большую роль может иметь возрастной отбор, закономерности которого мы обсуждали в начале этой главы. Возможность быстрого преобразования генетической структуры популяций под воздействием возрастного отбора усиливает их временную генетическую дивергенцию и именно поэтому поддерживает генетическую структуру популяции в целом, способствует ее быстрому направленному преобразованию, не говоря уже о непрерывном обогащении общего генофонда.
Развиваемые здесь взгляды, как нам кажется, находятся в столь же хорошем соответствии с данными экспериментальной и популяционной генетики, что и основные постулаты синтетической теории эволюции, но в значительно большей мере учитывают достижения современной экологии. Можно поэтому считать важнейшей задачей эволюционной экологии изучение динамики численности и динамики генетической структуры микропопуляций, являющихся частью биологических макросистем более высокого ранга. Общие выводы из развиваемых здесь взглядов существенно отличаются от главных выводов синтетической теории эволюции.
Быстрое эволюционное преобразование популяций (микроэволюция) не требует разделения видовой популяции па частично изолированные популяции средних размеров (один из важнейших постулатов теории Майра—Райта; также см. гл. II). Никакой изоляции микропопуляций, помимо той, которая определяется это логическими механизмами и разной динамикой численности, не требуется. Наоборот, постоянный обмен генами ускоряет процесс эволюционного развития. Биотопическое разнообразие занимаемой популяцией территории ускоряет микроэволюционный про-
{211}
цесс, но и оно не является необходимым, так как даже ничтожное изменение условий среды или случайное нарушение структуры популяции в начале сезона размножения животных приведет к изменению типа динамики численности со всеми вытекающими отсюда последствиями.
Таким образом, снимается ряд трудностей и одно из главных противоречий синтетической теории эволюции, согласно которым наиболее быстро эволюционируют изолированные популяции. Между тем любому беспристрастному исследователю ясно, что магистральный путь эволюции проходит не в условиях изоляции, а на больших участках арены жизни, открытых всем ветрам борьбы за существование. Вся история животного мира ясно показывает, что эволюция — это тот процесс, который привел от амебы к человеку, и что магистральная линия преобразований заключается в приспособлении к макросреде — к жизни на суше, в лесу, степи, на горах. Нам могут возразить, что особо быстрые преобразования видов действительно совершаются на островах. Однако островные формы — это «монстры эволюции». В том-то и дело, что эволюция на больших участках арены жизни происходит не медленнее, чем в условиях островной изоляции, но она не допускает уклонений с магистрального пути и поэтому не дает столь бросающихся в глаза форм, как дронт, дарвиновы вьюрки, моа, галапагосские игуаны и т. п. Необходимо, однако, отметить, что и на островах главным фактором эволюции является естественный отбор, а не генетико-автоматические процессы, основанные на принципе основателя, хотя именно здесь они могут иметь относительно большое значение. Представление о доминирующей роли случайности в эволюции островных форм возникло просто потому, что «монстры эволюции» резче бросаются в глаза. Тщательные исследования, проведенные в новейшее время [Radovanovic, 1959, 1961; Corbet, 1961, 1964; Berry, 1964; Foster, 1964; Eisentraut, 1965; Soule, 1966, и др.], показали, что, как правило, и на островах эволюция подчиняется тем же закономерностям, что и на материках, а общие особенности островных форм объясняются общими особенностями островов, как арены жизни (обедненность биоценозов, малочисленность хищников и т. п.). Обедненность островных биоценозов и как следствие этого ослабление борьбы за существование допускают известные отклонения от магистральной линии эволюции и, увеличивая роль случайных факторов, могут привести к ускорению темпов эволюционных преобразований. При этом не следует забывать, что в иных случаях увеличение темпов эволюции может быть связано как раз с обогащением биоценозов. Простейший случай — введение в биоценоз нового хищника повышает интенсивность естественного отбора со всеми вытекающими отсюда последствиями. Более сложную ситуацию иллюстрируют экспериментальные исследования Пайментэла [Pimentel, 1965], который показал, что совместное обитание двух близких видов не только приводит
{212}
к их быстрому эволюционному преобразованию, но и к эволюции кормовых растений.
Высказанные соображения вытекают из теоретического анализа, основанного на сопоставлении данных популяционной генетики с данными популяционной экологии. Эти соображения находят себе фактическую аргументацию в общем ходе эволюционного процесса.
Если бы темпы внутривидового формообразования на сплошном участке ареала вида определялись бы подразделением видовой популяции на частично изолированные локальные популяции, дифференциация которых происходит под совместным влиянием случайных процессов (принцип основателя) и индивидуального и межпопуляционного отбора, как это вытекает из схемы Райта и как на этом единодушно настаивают сторонники синтетической теории эволюции, то в пределах отдельных ландшафтных зон должны были бы наблюдаться существенные различия между разными группами животных. Мелкие виды, для которых река — уже преграда, образуют множество локальных популяций, которых крупные, широко мигрирующие виды почти не образуют. Тем не менее темп эволюционных преобразований крупных форм существенно не отличается от темпа преобразования мелких. Это противоречие со схемой Райта осталось, как нам кажется, незамеченным.
Воспользуемся для анализа зоной тундры. На ее необъятной территории можно насчитать сотни и тысячи локальных популяций леммингов, узкочерепной полевки, полевки Миддендорфа, полевки-экономки, красной полевки, землероек. Несмотря на это, мелкие млекопитающие тундры поразительно однообразны, они почти не образуют групп, которые бы заслуживали выделения в самостоятельные внутривидовые таксоны [Rausch, 1953; Шварц, 1963]. Крупные млекопитающие тундры (северный олень, песец) локальных популяций фактически почти нигде (за исключением островов) не образуют. Этому препятствуют миграции на громадное расстояние. Тем не менее внутривидовая дифференциация этих видов не меньше, чем у арктических полевок. В противоположность полевкам крупные арктические виды образуют несколько четко выраженных подвидов [Друри, 1949; Rausch, 1953; Herre, 1964]. Сказанное не означает, что формообразование у арктических млекопитающих идет медленно. Есть основания утверждать, что оно идет с большой скоростью, но не сопровождается дивергенцией популяций: вид на громадном пространстве развивается как единое целое. В этом отношении особенно показателен пример с леммингами (Lemmus, Dicrostonyx) — это один из самых молодых родов, но степень их морфофизиологического своеобразия исключительна.
Наоборот, виды, которые буквально олицетворяют собой схему Райта, эволюционируют крайне медленно. Лучший пример — уральская пищуха. Н. С. Гашевым в нашей лаборатории обследо-
{213}
ваны популяции пищухи (Ochotona hyperborea) из пяти пунктов на Полярном Урале. Расстояние между популяциями от 10 до 60 км. Учитывая горный рельеф района работ, исследованные популяции можно считать развивающимися самостоятельно, хотя незначительный обмен особями между ними должен происходить, так как перемещения пищух на несколько километров (в течение одного сезона) доказаны [Гашев, 1966]. Это типичные небольшие, частично изолированные популяции. Их изучение помогает, следовательно, составить себе представление о том, как работает схема Райта (см. главу III) в природе. Н. С. Гашев изучил 16 морфологических признаков (вес тела, пропорции тела и серию других показателей) этих популяций. Они оказались идентичными (это подтверждено статистической обработкой материала). Мы приводим лишь показатели отдельных популяций по наиболее важным таксономическим признакам. Кондилобазальная длина черепа у сравниваемых популяции была равной: 36,3; 35,4; 35,4; 35,3; 35,3; 35,4 мм; длина зубного ряда: 7,6; 7,3; 7,1; 7,2 мм; скуловая ширина: 19,6; 19,6; 19,5; 19,5; 19,6 мм; длина носовых костей: 11,4; 11,3; 11,4; 11,1; 11,1 и мм; индекс длины черепа (по соотношению к длине тела): 106,9; 106,2; 106,3; 106,4; 106,6%о; индекс длины зубного ряда (по отношению к кондилобазальной длине): 20,7; 20,9; 20,9; 20,3; 20,3%; индекс скуловой ширины: 54,1; 55,3; 55,1; 55,3; 55,3%. По всем без исключения таксономически существенным признакам сравниваемые популяции оказались тождественными (приходится удивляться, что за счет случайностей при сборе материала между популяциями практически не обнаружены даже несущественные различия).
По морфофизиологическим показателям между популяциями также обнаружены лишь ничтожные различия. Это иллюстрируется следующими примерами: индекс сердца: 5,6; 4,1; 4,6; 4,7; 4,5%о; индекс печени: 49,3; 42,1; 48,4; 50,6; 49,1‰; индекс почки: 7,3; 7,5; 6,9; 6,9; 7,2‰; индекс надпочечника: 0,11; 0,17; 0,16; 0,15; 0,16‰; индекс легких: нет данных; 9,1; 9,6; 9,1; 7,7‰.
Развиваясь в условиях изоляции в течение длительного времени, популяции пищухи не подвергались дивергенции, так как развитие их проходит в одинаковых условиях существования.
Другой пример: дифференциация популяции северных экономок (Microtus oeconomus), которые отличаются от южных по относительной длине хвоста, относительной длине задней ступни, индексу кондилобазальной длины черепа, ширине межглазничного промежутка и скуловой ширине. О. А. Пястолова [1967] провела сравнение по этим показателям трех изолированных популяций полевок (Ямал, Полярный Урал, острова в низовьях Оби). Сравнивались животные одинакового возраста и размеров, что значительно повышает достоверность результатов. Получены следующие данные. Длина хвоста в соответствующих популяциях оказалась равной (указаны средние величины): 38,5; 40,0; 38,2 мм; длина ступни: 19,1; 18,5; 19,4 мм; кондилобазальная длина чере-
{214}
па: 26,2; 26,4; 26,6 мм; скуловая ширина: 14,2; 14,2; 14,0 мм; ширина межглазничного промежутка: 3,6; 3,5; 3,5 мм. И в данном случае изолированные, но развивающиеся в одинаковых условиях популяции оказались практически тождественными.
В нашей лаборатории В. Н. Большаков изучал популяции прометеевой полевки (Prometheomys schaposhnikovi) на Главном Кавказском хребте (18 особей) и Аджаро-Имеретинском хребте (72 особи). Изоляция этих популяций полная. Исследования показали практически полное сходство животных этих популяций по размерам тела и краниологическим показателям. Длина тела равна соответственно 130,6 и 126,9 мм (сравниваются одновозрастные зверьки), кондилобазальная длина черепа — 30,62 и 30,53 мм, длина зубного ряда — 7,31 и 7,32 мм, скуловая ширина — 17,45 и 17,22 мм, высота мозговой части— 11,9 и 11,33 мм, длина лицевой части—18,43 и 18,51 мм, межглазничная ширина — 4,02 и 4,29 мм; длина диастемы — 9,03 и 8,99 мм. Очень сходными оказались сравниваемые популяции и по интерьерным признакам. Так как, что и следовало ожидать, различия здесь все же более значительны, мы приводим полученные средние величины со статистической ошибкой. Относительный вес сердца полевок с Главного Кавказского хребта равен 4,5±0,17‰, Аджаро-Имеретинского — 4,1±0,10; печени соответственно—49,3±1,71 и 50,4±0,95‰, почек — 5,4±0,18 и 4,5±0,11‰; относительная длина кишечника — 480±11 и 450±6; слепого отдела кишечника— 37±1 и 39±0,7‰.
Приводя многочисленные примеры быстрой эволюции на островах, сторонники синтетической теории эволюции нередко забывают указать и на факты совершенно иного рода. Так, тщательные исследования Флеминга [Fleming, 1962] показали, что возраст эндемичных родов Новой Зеландии неогеновый, видов — плейстоценовый. Но ведь новые роды материкового происхождения ничуть не старше (Lemmus, Dicrostonyx, возможно, Rangifer, Ovibos, Alces, Saiga, Passerina, Lagopus, Nyctea и др.).
Уже эти факты ясно показывают, что и на сплошном участке ареала вида нередко складываются условия, способствующие столь же быстрой эволюции, что и на островах, но в противоположность островным формам формы материковые характеризуются биологически существенными адаптивными особенностями, они находятся в основном русле эволюционного процесса. Нетрудно попять и первопричины их быстрой эволюции — это изменение условий среды. Все молодые роды позвоночных — обитатели тундры и тайги, наиболее молодых ландшафтных зон. Новые условия среды: вызвали вспышку формо- и видообразования 1. Невыгодная с точки зрения синтетической теории эволюции структура вида (слабо выраженное дробление вида на локальные популяции) не
1 И у растении в высоких широтах эволюция протекает с большей скоростью [Васильев, 1965].
{215}
снизила темп филогенетических преобразований северных форм. Причину этого мы видим в экологических механизмах эволюционного процесса.
Наконец, можно привести еще одно более общее доказательство в пользу развиваемых здесь взглядов. Если бы наличие локальных, генетически изолированных популяций было бы главной причиной, определяющей темп эволюционных преобразований, то при прочих равных условиях эволюция мелких оседлых форм происходила бы быстрее, чем крупных, способных к далеким миграциям, при которых широкий обмен генами неизбежен.
Темп эволюции лошадей существенно не отличается от темпа эволюции мелких степных грызунов. Быстрая смена видов характерна для эволюции слонов [Zeuner, 1955]. На это недавно обратил внимание Хехт [Hecht, 1965]. Упомянув о том, что, согласно теоретическим расчетам Холдена, для становления нового вида необходима смена не менее 300 000 поколений, Хехт указывает на палеонтологическом материале, что эта цифра завышена почти в 10 раз. Для эволюционного ряда Archidiscodon subplanifrons — A. planifrons — A. meridionalis — Elephas sp. потребовалась смена всего 100 000 поколений. Некоторые виды китов имеют возраст 15–45 тыс. лет [Davies, 1963]. Вспомним, наконец, эволюцию человека: за несколько сот тысячелетий пройден путь от питекантропа до современного человека, возникло по крайней мере несколько эволюционных тупиков родового ранга. Вряд ли наши предки образовывали локальные популяции, а способность их к миграциям кажется сейчас доказанной. Подобные факты, а они отнюдь не единичны, заставляют признать возможность быстрой эволюции на сплошном участке ареала вида. Выдвигаемая нами схема дает им естественное объяснение. Анализ вероятного значения динамики пространственной структуры популяций приводит нас к заключению, что на сплошном участке ареала вида эволюция может происходить с большой скоростью, не сворачивая с магистрального пути, определяемого условиями среды и соответствующим направлением естественного отбора. Главная движущая сила процесса — экологические механизмы преобразования популяций.