
Проблемы Эволюции |
О взрывном видообразовании, протекающем на наших глазах в высыхающем Аральском море. Омск: Изд-во Омского государственного педагогического университета, 2003. 382 с.
Глава 1. ИСТОРИЯ ИЗУЧЕНИЯ ДВУСТВОРЧАТЫХ МОЛЛЮСКОВ АРАЛЬСКОГО МОРЯ. МАТЕРИАЛ И МЕТОДИКА
Начальные сведения по фауне Аральского моря были получены попутно с другими исследованиями в период дипломатического и торгового проникновения России в Среднюю Азию. Так, по публикациям девятнадцатого – начала двадцатого века в общем был выявлен видовой состав двустворчатых моллюсков. Для Аральского моря были указаны: Dreissensia (=Dreissena) polymorpha var. aralensis Andr., Dreissensia polymorpha var. obtusecarinata Andr., Dreissensia pallasi Andr., Dreissensia caspia Eichw., Adacna (=Hypanis) minima Ostr., Adacna vitrea Eichw., Adacna laeviuscula Eichw., Cardium (=Cerastoderma) edule L. (Basiner, 1848; Мартенс, 1874; Аленицын, 1876; Андрусов, 1897; Остроумов, 1907; Берг, 1908).
В обстоятельной сводке С.А. Сидорова (1929) приведено для Аральского моря и его ближайших окрестностей 37 видов моллюсков, в том числе 13 двустворок, обитавших непосредственно в море: Dreissensia polymorpha Pall., D. polymorpha var. aralensis Andr., D. polymorpha var. obtusecarinata Andr., D. pallasi Andr., D. caspia Eichw., D. rostriformis Desh., Adacna minima Ostr. forma a и forma b, A. vitrea Eichw., Cardium edule L., C. edule var. lamarcki Reev, C. edule var. lamarcki Reev (?).
В определителе моллюсков Советского Союза В.И Жадиным (1952) для Аральского моря приведены Dreissena polymorpha var. aralensis (Andr.), D. polymorpha var. obtusecarinata (Andr.), D. pallasi (Andr.) и D. caspia (Eichw.), Adacna vitrea Eichw., A. laeviuscula Eichw., Cardium edule L.
Исследование животного мира Арала, в основном, как и во время нашей работы на Аральском море, носило чисто прикладной рыбохозяйственный характер и сводилось, за редким исключением, к подсчету численности и биомассы отдельных групп организмов и изучению их распределения по акватории моря в лучшем случае с приведением списка систематических групп, в котором указаны вид или чаще род, без каких-либо описаний и иллюстраций, которые могли бы уточнить видовую принадлежность беспозвоночных (Никитинский, 1933; Бенинг, 1934, 1935; Деньгина, 1957, 1959а, б; Яблонская, 1960; Гаврилов, 1967, 1968, 1972, Кортунова, 1967, 1970; и др.).
Работы по биологии и солеустойчивости аральских беспозвоночных (Карпевич, 1964, 1975; Хусаинова, 1958; Бекмурзаев, 1970; и др.) также не сопровождались видовым описанием и рисунками животных.
Существенным вкладом в изучение фауны Аральского моря явилось издание «Атласа беспозвоночных Аральского моря» (1974), обобщающего всю информацию по фауне беспозвоночных Арала, имевшуюся к началу 1970-х годов, в котором по результатам ревизии аральских моллюсков Я.И. Старобогатовым указано 9 видов и подвидов двустворок: Hypanis vitrea bergi Star., H. minima sidorovi Star., H. minima minima (Ostr.), Dreissena polymorpha aralensis (Andr.), D. p. obtusecarinata (Andr.), D. caspia pallasi (Andr.), Cerastoderma lamarcki lamarcki (Reeve), Cerastoderma umbonatum (Wood), Abra (=Syndosmya) ovata (Phil.).
Изменчивость аральских двустворок, как, впрочем, и большинства других групп животного мира этого водоема, практически не исследовалась.
Единственным источником обширной информации по изменчивости аральских Cerastoderma являются работы М. Бетсона (Bateson, 1889, 1890). Автором исследовался ряд террас соленых озер (Чумышкуль, Джаман-клыч, Джаксы-клыч) по северо-восточному побережью Аральского моря, содержащих раковины Cerastoderma (Cardium edule в оригинале), при этом было отмечено, что от верхних террас к нижним изменяется форма, толщина и интенсивность окрашивания раковины Cerastoderma. У моллюсков удлиняется задняя часть раковины, происходит уплощение раковины и уменьшаются размеры раковин и число ребер с 18–22 на первой террасе до 14–15 на седьмой. Подобные изменения характерны и для Cerastoderma (Cardium edule в оригинале) из соленых озер Египта (Bateson, 1889).
В обстоятельном труде С.А. Сидорова (1929) при описании видов аральских Dreissena основное внимание уделено фактам нахождения вида и описанию систематических отличий. По каждому виду дрейссен даны промеры нескольких экземпляров. Только для Dreissena polymorpha из оз. Акчатау (низовья реки Сырдарьи) проанализирован полиморфизм по контуру створок и положению киля. При обсуждении видового состава аральских Hypanis размеры моллюсков приведены без указания вида, для Cerastoderma дано число ребер и отношение ширины раковины к ее длине.
Н.З. Хусаинова (1958), описывая биологические особенности массовых донных беспозвоночных Арала, не привела конкретных данных по изменчивости аральских двустворчатых моллюсков за исключением общих замечаний. Так, для аральских Hypanis дано морфологическое описание без подразделения материала на виды. Для Cerastoderma отмечены две формы раковин, различающиеся вытянутостью заднего края, количеством и шириной ребер, положением макушек, отношением высоты раковины к длине и высоты к толщине без приведения цифрового материала. Изучая фауну култуков восточного побережья, Н.З. Хусаинова (1958, 1960) замечает в отношении Cerastoderma, что при солености 16–23 ‰ особи мельче, чем в собственно Арале; при перенаселении моллюсками некоторых участков дна отмечались несимметричные створки раковин, либо наличие уродливых раковин с вмятинами и искривлениями.
Я.И. Старобогатовым (1974) при описании видов Dreissena, Hypanis и Syndosmya в «Атласе беспозвоночных Аральского моря» указаны колебания основных морфометрических индексов.
Анализируя публикации по изменчивости двустворчатых моллюсков, следует отметить, что информация довольно скудна, как по изменчивости переживших экологический кризис аральских Cerastoderma и Syndosmya, так и по изменчивости Cerastoderma из других южных морей России. Из доступных литературных источников это работы О.А. Гримма (1876), указывающего количество ребер для каспийских Cerastoderma.
Полиморфизм Cardium (=Cerastoderma) edule L. привлекал внимание исследователей с конца XIX века. На основании различий в контурах створок раковин, числе ребер, строении и относительных размерах лигамента, окраске и прочих признаках неоднократно предпринимались попытки разделения этого вида на подвиды и, в последующем, на виды (Vinsent, 1882; Милашевич, 1916; Chavan, 1945; Mars, 1949; Petersen, 1958; Grossu, 1961; Boyden, 1973; Brock, 1978; Бессонов с соавт., 1970; Скарлато, Старобогатов, 1972; и др.). Однако данных по морфометрическому анализу раковин Cerastoderma в указанных источниках практически не обнаружено.
Только по Cerastoderma glaucum Poiret из Черного моря найдено исследование Т.В. Михайловой (1985), которая приводит морфометрические индексы, констатируя, что отношения линейных размеров раковин почти не зависят от возраста моллюсков и условий обитания и могут быть использованы как видовой признак.
Скудность информации по изменчивости основных морфологических параметров видов рода Cerastoderma, вероятно, обусловлена довольно стабильными условиями обитания этих моллюсков в стабильных насыщенных экосистемах, когда все отклонения жестко отметаются отбором. Невысокая вариабельность морфологических признаков переключает внимание исследователей на изменчивость Cerastoderma на физиологическом, биохимическом и генетическом уровнях (например: Brock, 1978, 1987, 1989; и др.).
Сведений по эволюционным преобразованиям современных двустворчатых моллюсков Аральского моря в научной литературе не обнаружено, информация о публикациях по генезису двустворок приведена в соответствующих разделах работы.
Полиморфизм акклиматизированного в Арале двустворчатого моллюска Syndosmya segmentum был отмечен еще в конце 1970-х годов (Андреева, 1980). Проблема изменчивости аральских двустворок вновь четко обозначилась в начале 1990-х годов. После возобновления в 1989 г. исследований экосистемы Аральского моря Казахским НИИ рыбного хозяйства нами (Андреев, Андреева, 1995б; Андреева, 1996) было обращено внимание на крайний полиморфизм раковин Cerastoderma, содержащихся в пробах бентоса. В последней экспедиции на Аральское море в 1994 г. Н.И. Андреевым были проведены специальные сборы с целью изучения изменчивости раковин аральских двустворчатых моллюсков. Опираясь на эти сборы, а также на сохранившиеся количественные пробы бентоса стандартных съемок Аральского моря, сделана попытка реконструкции эволюционных преобразований его двустворчатых моллюсков в условиях современного экологического кризиса.
В общей сложности наблюдения за видовым составом и изменчивостью двустворчатых моллюсков Аральского моря с большими перерывами охватывают период с 1900 по 1994 гг. Для этих целей использованы коллекции: сборы экспедиции Л.С. Берга (1900–1902 гг.), хранящиеся в фондах Зоологического института (ЗИН) РАН; сохранившиеся пробы бентоса весенней стандартной съемки 1965 г. из хранилища Аральского отделения Казахского НИИ рыбного хозяйства (КазНИИРХ), обширные сборы авторов (1976–1982, 1989–1994 гг.) в рейсах по стандартной сетке станций Аральского отделения
КазНИИРХ и в береговых и култучных рейсах (рис. 1.1), а также сборы Cerastoderma 1992 г. из залива Тщебас Большого моря, любезно предоставленные А.А. Филипповым.
Динамика численности и биомассы двустворчатых моллюсков и зообентоса в целом до 1963 г. приводится по литературным данным, с 1963 по 1975 гг. по картотеке первичной обработки бентоса из фондов Аральского отделения КазНИИРХ, а с 1976 по 1994 гг. по собственным сборам. В период падения уровня моря и осолонения его вод гидробиологические сборы проводились Аральским отделением
КазНИИРХ ежегодно по постоянной сетке станций, принятой в 1959 г. (рис. 1.1). Располагались станции равномерно по акватории моря от глубины 2.5 и до максимальной. Количество станций в связи с падением уровня моря постепенно уменьшилось со 134 до 54 в 1981 г., когда была в последний раз проведена одновременная съемка всего моря. В 1982 и 1993–1994 гг. проводились съемки только Малого моря, а с 1989 по 1992 гг. в совместных экспедициях с сотрудниками Государственного океанографического института (ГОИН) проведены съемки и Большого моря. Одновременно с отбором гидробиологических проб выполнялись стандартные гидролого-гидрохимические исследования. Сбор и обработка зообентоса проводилась все годы по стандартной методике (Инструкция …, 1930; Яблонская, 1960, Андреева, 1989; Андреев, Андреева, 1997а). Всего было обработано 756 проб стандартных и прибрежных рейсов 1976–1982, 1989–1994 гг. и проанализировано 2380 карточек первичной обработки бентоса за 1963–1975 гг. из фондов Аральского отделения КазНИИРХ.
<![if !vml]>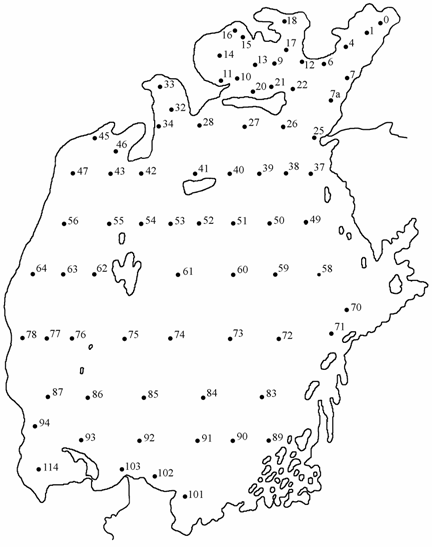 <![endif]>
<![endif]>
Рис. 1.1. Расположение станций сбора зообентоса на Аральском море.
Для контрольных выборок использованы массовые сборы моллюсков из различных районов Аральского моря за 1976–1994 гг. (рис. 1.2), а также уцелевшие коллекции экспедиции Л.С. Берга и стандартной съемки 1965 г.
Схемы количественного распределения моллюсков составлены методом прямолинейной интерполяции.
Для сравнения изменчивости Cerastoderma isthmicum и Syndosmya segmentum из Аральского моря и из других водоемов, а также изменчивости внутри видов рода Cerastoderma привлечены сборы из северной части Каспийского моря за 1892, 1897 гг. и сборы экспедиции С.А. Зернова из Черного моря у берегов Болгарии и Румынии за 1911 г. из коллекций ЗИН РАН.
При характеристике раковин моллюсков руководствовались общепринятой терминологией, используемой при изучении морфологии двустворок (Скарлато с соавт., 1990). Исследовались форма раковины, ее окраска, величина апикального угла, степень погружения раковины в грунт, строение замка и отдельных его частей, форма и размеры лигамента и мускулов-замыкателей, конфигурация и ширина мантийной линии, а также ряд счетных и пластических признаков, описываемых ниже.
Для характеристики изменчивости раковин Cerastoderma подсчитывалось общее количество ребер, покрывающих раковину, включая слабовозвышающиеся (сглаженные) и еле заметные в виде отпечатка (следовые) ребра, иногда заметные только при изменении освещения, а также количество нормально развитых (возвышающихся) ребер.
<![if !vml]>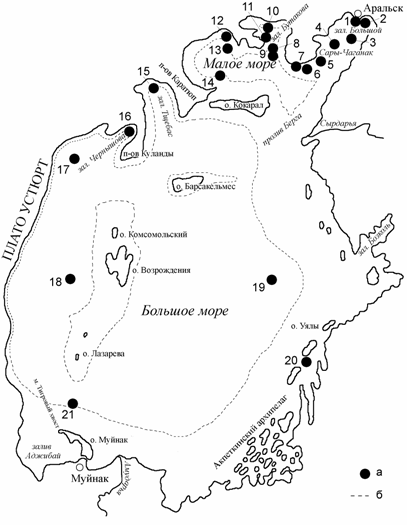 <![endif]>
<![endif]>
Рис. 1.2. Схема станций контрольных выборок.
а – станции: 1 – Аральская бухта; 2 – прибрежье залива Сары-Чаганак; 3 – станция 0; 4 – станция 1; 5 – станция 4; 6 – станция 6; 7, 11 – береговые выбросы; 8 – станция 17; 9 – вход в залив Бутакова; 10 – залив Бутакова; 12 – станция 16; 13 – станция 15; 14 – район станции 11; 15 – залив Тщебас; 16 – залив Чернышова; 17 – станция 47; 18 – станция 63; 19 – район станции 59; 20 – район острова Толмачева; 21 район станции 93. б – примерное положение береговой линии в августе 1992 г.
Во второй трети раковин церастодермы нормально развитые ребра обычно имеют одно или два отличающихся по выраженности (высоте, иногда и ширине), ярко окрашенных с разноцветными пятнами ребра. В редких случаях их может не быть. Положение этих ребер, именуемых в дальнейшем «более выраженное ребро (ребра)», значительно варьирует в популяциях аральских сердцевидок и может служить в качестве одного из элементов генетической характеристики популяции. Ребра аральских церастодерм обычно в передней трети раковины покрыты поперечными известковыми чешуйками, в сборах 1989–1994 гг. на створках наряду с чешуйками заметны довольно выступающие конхиолиновые складочки, положение которых весьма изменчиво (рис. 1.3).
<![if !vml]>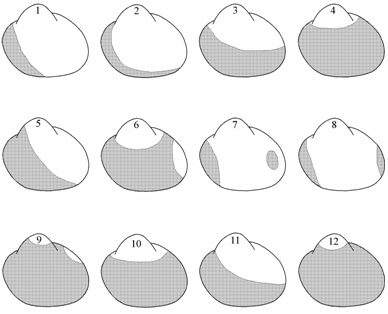 <![endif]>
<![endif]>
Рис. 1.3. Схема положения чешуек и конхиолиновых складочек на створках Cerastoderma Аральского моря (по: Андреева, Андреев, 2000а). Цифрами обозначены типы расположения чешуек и конхиолиновых складочек.
При анализе изменчивости лигамента выделялись такие отделы: выпуклая часть лигамента, расположенная позади макушек, ее хвостообразное продолжение и, иногда, впереди макушек имелось образование в виде продолжения лигамента (передний лигамент). И переднее продолжение лигамента, и хвостообразный придаток обычно имеют отличную от выпуклой части структуру: они представлены обычно слоем слияния (Allen, 1960; и др.) и состоят только из конхиолина, окрашенного в желтый или коричневый всех оттенков янтаря цвет. Выпуклая часть имеет типичное для двустворок строение (Скарлато с соавт., 1990), причем ламеллярный слой просвечивает отдельными полосками, создавая наряду с расширениями и сужениями членистость лигамента.
Изменчивость апикального угла изучалась по рисункам раковин в стандартном положении, полученных при помощи рисовального аппарата (рис. 1.4).
<![if !vml]>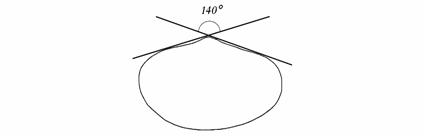 <![endif]>
<![endif]>
Рис. 1.4. Схема измерения апикального угла двустворок.
Замки аральских Cerastoderma и Syndosmya чрезвычайно разнообразны, в типичном случае (рис. 1.5) у Cerastoderma имеется шесть зубов (два передних латеральных, два кардинальных, два задних латеральных) на правой створке и четыре зуба (передний латеральный, два кардинальных и задний латеральный) на левой, у S. segmentum – четыре на правой створке (передний латеральный, два кардинальных и задний латеральный) и один кардинальный на левой.
Глубина погружения раковины в грунт (рис. 1.6) определялась по положению четко выраженной границы обрастаний на части раковины, находящейся вне грунта (1976–1994 гг.), и по следу, оставленному грунтом (1989–1994 гг.) в передней части, погруженной в грунт. В случае погружения раковины в грунт полностью она не несет следов обрастаний. След от погружения раковины в грунт хорошо просматривается лишь на экземплярах длиною более 12–15 мм, более мелкие экземпляры еще не имеют достаточно хорошо развитых обрастаний и не включались в анализ по данному признаку.
<![if !vml]>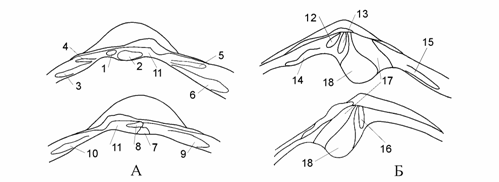 <![endif]>
<![endif]>
Рис. 1.5. Замок.
На А – Cerastoderma: вверху зубы правой створки — кардинальные: 1 – передний (первый), 2 – задний (второй); латеральные: 3 – первый передний, 4 – второй передний, 5 – первый задний, 6 – второй задний. Внизу зубы левой створки — кардинальные: 7 – передний (первый), 8 – задний (второй); латеральные: 9 – передний, 10 – задний. 11 – нимфа. На Б – Syndosmya: вверху зубы правой створки — кардинальные: 12 – передний (первый), 13 – задний (второй); латеральные: 14 – передний, 15 – задний. Внизу зубы левой створки: 16 – кардинальный. 17 – нимфа, 18 – ямка внутреннего лигамента.
При анализе формы раковины двустворок использован ряд индексов, вычисленных на основе стандартных и дополнительных промеров (рис. 1.7).
При помощи окуляр-микрометра микроскопа МБС-9 измерялись высота раковины (Н), длина раковины (L), выпуклость раковины (В), длина макушек (а), длина лигамента (с), длина выпуклой части лигамента (d), и расстояние (l1), измеренное по продольной оси раковины от наиболее выпуклой точки переднего края до центрального ребра, проходящего через середину макушек, а также расстояние (l2), измеренное от перпендикуляра, опущенного из середины макушки на прямую, соединяющую наиболее удаленные точки переднего и заднего края раковины, до наиболее выпуклой точки переднего края раковины.
<![if !vml]>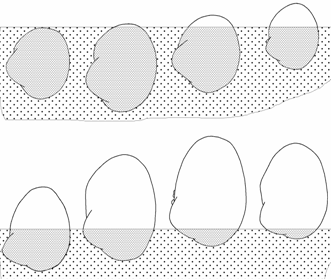 <![endif]>
<![endif]>
Рис. 1.6. Схема выхода из грунта и смещения макушек к переднему концу раковины у Cerastoderma Аральского моря.
В качестве индексов использованы отношения высоты раковины к ее длине (Н/L), выпуклости раковины к ее длине (В/L), выпуклости раковины к ее высоте (В/Н), длины макушек к длине раковины (а/L), длины макушек к длине лигамента (а/с), длины макушек к длине выступающей части лигамента (а/d), расстояние от переднего края раковины до центрального ребра к длине раковины (l1/L), а также положение середины макушек по отношению к переднему краю раковины – (l2/L). Индекс В/Н является видоспецифическим для ряда двустворчатых моллюсков (Наумов, 1976), по индексу В/L разделены азово-черноморские виды церастодерма (Скарлато, Старобогатов, 1972).
Промеры раковин и рисунки сделаны при помощи микроскопа МБС-9 и модернизированного рисовального аппарата. Сравнение кривизны фронтального сечения створок раковин Cerastoderma проведено компараторным методом Я.И. Старобогатова (Логвиненко, Старобогатов, 1971; Скарлато с соавт., 1990).
<![if !vml]>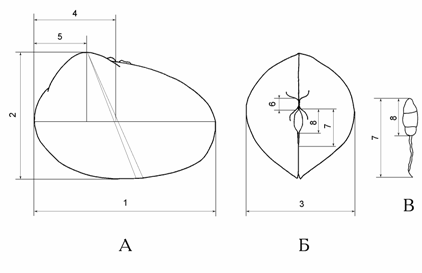 <![endif]>
<![endif]>
Рис. 1.7. Стандартные промеры раковин.
А – вид спереди, Б – вид сверху, В – лигамент: 1 –длина раковины (L), 2 – высота раковины (H), 3 – выпуклость раковины (В), 4 – расстояние от переднего края до центрального ребра (l1), 5 – расстояние от переднего края раковины до середины макушек (l2), 6 – длина макушек (a), 7 – длина лигамента (с), 8 – длина выпуклой части лигамента (d).
Таким образом, характеристика раковин аральских Cerastoderma проводилась по 26 признакам, Syndosmya по 16, из которых наиболее значимыми для Cerastoderma за весь период исследований оказались 20: восемь морфометрических индексов, форма контура створки, степень погружения раковины в грунт, количество ребер (общее и нормально развитых), форма лигамента, строение замка, окраска раковины снаружи и изнутри, положение более выраженного, ярко окрашенного ребра (ребер), симметрия створок, наличие зияния, ширина мантийной линии, для Syndosmya – 5: форма створки и лигамента, величина апикального угла, степень изгиба замочной площадки, конфигурация мантийной линии. Анализу было подвергнуто более 1000 экз. Cerastoderma, 1400 экз. Syndosmya из дночерпательных проб ряда станций стандартной сетки за 1965, 1976–1982, 1989–1994 гг., дополнительных сборов, и из коллекций ЗИН РАН (количественные сборы по Cerastoderma 1990 и 1994 гг. обработаны полностью). В целом же за весь период исследований просмотрено около 7500 экз. Cerastoderma и 44 тыс. экз. Syndosmya в количественных пробах стандартной съемки моря и прибрежных рейсов, 3270 экз. при экспериментальных работах по солеустойчивости аральских Cerastoderma, Syndosmya и Dreissena, а также большое количество раковин из береговых выбросов в различных участках побережья Арала. Объем просмотренного материала по Dreissena и Hypanis из количественных сборов много меньше: 985 и 41 экз. соответственно.