
Проблемы Эволюции |
О взрывном видообразовании, протекающем на наших глазах в высыхающем Аральском море. Омск: Изд-во Омского государственного педагогического университета, 2003. 382 с.
При описании среды обитания во внимание приняты только наиболее значимые для жизнедеятельности двустворчатых моллюсков параметры экосистемы Аральского моря. Желающих подробно ознакомиться с произошедшими изменениями и современным состоянием экосистемы Аральского моря отсылаем к специализированным сводкам: «Гидрометеорология и гидрохимия морей СССР: т. 7. Аральское море» (1990) и «Гидрофауна Аральского моря в условиях экологического кризиса» Н.И. Андреева (1999).
Аральское море – своеобразный водоем. Его главная отличительная черта – отсутствие связи с Мировым океаном, которой не было и в геологическом прошлом моря (Косарев, 1975). Расположено Аральское море на территории двух государств: юго-западная часть лежит в пределах республики Узбекистан, северо-восточная – республики Казахстан.
Согласно О.М. Житомирской (1964), район Аральского моря занимает крайнее северное положение в зоне континентального субтропического климата. Эта территория получает большое количество солнечного тепла. В среднем за год суммарная радиация составляет здесь 5860 МДж/м2. Радиационные особенности и положение в глубине обширного азиатского материка вызывают засушливость и континентальность климата.
Атмосферных осадков за год выпадает 100–115 мм. Максимальная температура воздуха летом +43°, минимальная — зимой –36°.
Исходя из целей работы, при описании гидролого-гидрохимического режима Аральского моря был дан режим моря до начала интенсивного водопотребления, отражающий условия существования флоры и фауны до зарегулирования стока рек, и показаны его изменения во время сбора гидробиологического материала для данной работы (по 1994 г. включительно). Период до 1961 г. считается периодом квазистационарного режима Аральского моря.
По данным Р.В. Николаевой (1969), составившей морфометрическое описание Аральского моря при его среднемноголетнем уровне (53.00 м абс.), площадь моря, включая острова, была 68320.5 км2; наибольшая глубина – 69 м, средняя – 16.1 м; площадь водной поверхности – 66085.6 км2; объем водных масс – 1061.6 км3. Длина береговой линии (без островов) превышала 4430 км. Наибольшая длина моря была 424 км, ширина – 292 км. Общее число островов на Арале площадью более 0.01 км2 составляло около 1100 с общей площадью 2234.9 км2, наиболее крупные острова – Кокарал (311 км2) и Барсакельмес (170 км2).
По строению котловины и некоторым особенностям гидрологического режима Аральское море делят на две части: северо-восточную, отделенную островом Кокарал, называемую Малым морем, и все, что лежит южнее острова Кокарал под названием Большое море. В свою очередь Большое море разделяет подводная гряда от южной оконечности полуострова Куланды через острова Комсомольский, Возрождения, Константин, Лазарева к острову Муйнак. К западу от гряды расположены наиболее глубоководные районы моря, к востоку – большая по площади асимметричная котловина, вытянутая в северном направлении (рис. 3.1).
В юго-восточной части Аральского моря располагался своеобразный Акпеткинский архипелаг, включающий более 500 островов и многочисленные заливы-култуки.
Анализ многолетних колебаний уровня (Львов, 1959, 1964) показал, что в период 1911–1961 гг. средняя многолетняя высота уровня над нулем поста Аральской ГМО была 152.6 см, или 53.06 м над уровнем океана в Балтийской системе отметок. Уровень моря изменялся от года к году с определенной цикличностью, равной 10–11 годам и совпадающей с цикличностью солнечной активности. Амплитуда этих многолетних колебаний уровня не превышала 1 м. При многолетних колебаниях уровня площадь и объем вод Аральского моря изменялись примерно на 5 %, соленость воды практически не изменялась. Помимо многолетних колебаний уровня, Аралу были свойственны вековые колебания, периодичность которых равна примерно 60 годам, амплитуда — 3–4 м. При вековых колебаниях (прослежены с 1780 г.) площадь моря изменялась на 16 %, объем воды – на 17 %, а соленость могла изменяться на 3 ‰.
<![if !vml]>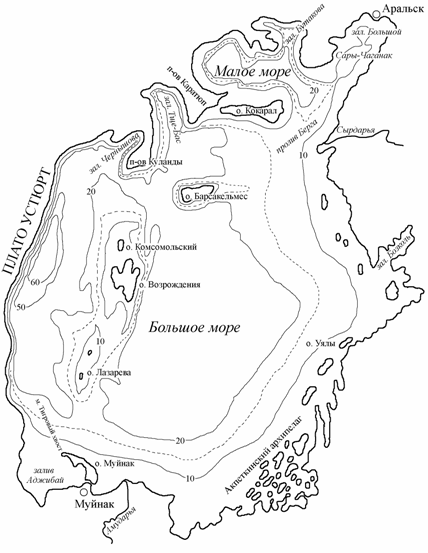 <![endif]>
<![endif]>
Рис. 3.1. Распределение глубин в Аральском море при отметке уровня 53 м абс. Пунктир – примерное положение береговой линии в августе 1992 г.
<![if !vml]>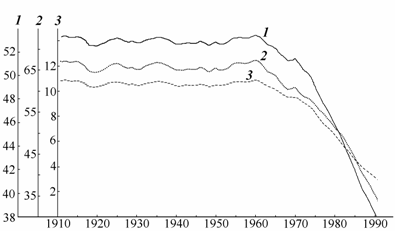 <![endif]>
<![endif]>
Рис. 3.2. Изменение уровня (1 – м абс.), площади (2 – ´103 км2) и объема (3 – ´102 км3) Аральского моря (по: Лобов с соавт., 1995).
С 1961 г., в связи с зарегулированием стока рек Амударьи и Сырдарьи, средний годовой уровень моря стал снижаться. Небольшое его повышение произошло лишь после экстремально многоводного 1969 г. (рис. 3.2).
Падение уровня моря заметно ускорилось с 1975 г., если за 1961–1974 гг. уровень понижался в среднем на 27 см в год, то за 1975–1985 гг. на 71 см в год (Каталог …, 1987; Гидрометеорология …, 1990). К концу 1989 г. уровень моря снизился до отметки 39.02 м абс., или на 14.4 м по сравнению с его значением за 1960 г. (Перминов с соавт., 1993).
В конце 1987 г. произошло расчленение Аральского моря на две практически обособленные части – Большое и Малое моря. В связи с большими поступлениями вод по р. Сырдарье в Малое море периодически из него происходил сброс воды и солей в Большое море, что по-видимому, привело к некоторому углублению дна в проливе Берга и снижению критической отметки, характеризующей условия разделения Большого и Малого морей до примерно 39.5 м абс. (Цыцарин, 1991; Цыцарин, Бортник, 1991). Если максимальный уровень Малого моря определялся критической отметкой 39.5 м абс., выше которой в нем не могли удерживаться воды, то уровень Большого моря продолжал снижаться и к моменту последней гидробиологической съемки в августе 1992 г. опустился до отметки 37.14 м абс.
В 1989–1992 гг. темпы снижения уровня моря несколько замедлились, что связано с многоводностью последних лет и значительным сокращением испаряющей поверхности.
Большое снижение уровня моря определило изменение морфометрии Арала. К 1993 г. осушилось 45 % его акватории (см. рис. 3.1.), на 70 % сократился объем и на 45 % уменьшилась средняя глубина. Остров Кокарал, разделяющий Малое и Большое моря, соединился с материком. Острова Комсомольский, Возрождения, Константин и обнажившаяся банка Бенинга соединились и образовали единый остров.
Небольшие острова по восточному побережью и Акпеткинский архипелаг оказались на суше и утратили географический смысл. В отдельных местах восточного побережья море отступило более чем на 80 км. Появился ряд новых островов на месте бывших банок. По мере снижения уровня наблюдался быстрый рост песчаных кос от острова Лазарева на юг в направлении бывшего мыса Тигровый хвост и на север к архипелагу Возрождения, а также от острова Барсакельмес в направлении восточного побережья, который привел к обособлению и интенсивному снижению водообмена между центральной частью Большого моря, северной его частью и глубоководной впадиной.
Берега Аральского моря в северо-восточном, северном и северо-западном районах характеризовались лопастной изрезанностью, побережья восточного и южного районов, сформировавшиеся в результате затопления пустынь, были сильно изрезаны (Лымарев, 1967). В результате отступления моря береговая линия восточного, юго-восточного и южного побережий Арала значительно выпрямилась, там полностью исчезли крупные мелководные заливы.
Водный баланс Аральского моря складывается из притока вод Амударьи и Сырдарьи, подземного притока, осадков на поверхности моря и испарения с поверхности моря. Приток вод к Аралу, практически, складывался из стока рек Сырдарьи и Амударьи, дававших 97 % всей воды, получаемой морем. Средний многолетний сток рек до 1961 г. был около 60 км3 в год (Асарин, 1973). Подземный приток, по последним данным, определен в размере 0.2 км3 в год и, обычно, не учитывается в водном балансе Арала (Асарин, 1975). Средняя многолетняя величина осадков на поверхности моря составляет около 130 мм в год, средний многолетний годовой слой испарения с поверхности моря 998 мм (Современный …, 1972; Бортник, 1990а).
До 1961 г. испарение с поверхности моря уравновешивалось поступлением вод Амударьи и Сырдарьи, что обеспечивало относительно стабильный его уровень. В связи с увеличением объемов безвозвратного изъятия стока рек с 1961 г. наблюдалась хорошо выраженная тенденция к увеличению отрицательных значений водного баланса и неуклонное падение уровня моря (Николаева, 1969; Современный …, 1972; Крицкий с соавт., 1973; Асарин, 1973, 1975; Косарев, 1975; Бортник, 1977а, б, 1978, 1980б, 1983; Micklin, 1988). К 1993 г. падение уровня моря составило почти 16 м.
При оценке естественного и антропического воздействия на многолетние изменения уровня моря для периода 1961–1985 гг. было установлено, что за счет естественных факторов уровень моря снизился на 2.7 м (23 % от фактического его падения). За счет же антропических факторов уровень моря снизился еще на 9.2 м (77 %). В результате интенсивного развития хозяйственной деятельности интегральный дефицит поступления речных вод в Аральское море достиг примерно 570 км3 (Бортник, 1990б).
Горизонтальную циркуляцию вод (течения) Аральского моря определяют главным образом ветры над морем и рельеф дна. Кроме того, определенное значение имеет конфигурация береговой линии, плотность вод и распространение речных вод в море. Установлено, что для Аральского моря характерны два основных типа циркуляций: антициклоническая при ветровых полях северных направлений и циклоническая при ветровых полях южных направлений (Симонов, 1954; Косарев, 1975).
Для вод Аральского моря характерен сильный прогрев летом и образование ледяного покрова зимой. Максимальная температура в поверхностных слоях воды (более 26°) наблюдалась в июле в юго-восточной и южной частях моря.
Значительный прогрев поверхностных слоев воды в летний период приводит к увеличению вертикальных градиентов температуры и формированию хорошо выраженного термоклина, верхняя граница которого в западной глубоководной котловине находится на глубине 10–15 м, нижняя — 25–30 м. В центральных районах существенные градиенты наблюдаются до дна. Температура воды придонного слоя воды на глубинах 50–60 м не превышает 3–4° (Гидрометеорология …, 1990).
Температурный режим Аральского моря в современный период стал более контрастным, произошло постепенное увеличение размаха внутригодовых колебаний температуры для всей толщи вод. Это связано как с постоянным понижением температуры замерзания морских вод по мере повышения ее солености, так и со значительным обмелением моря и уменьшением его теплозапаса. В результате, в зимний период практически вся толща вод выхолаживается до температуры замерзания, а летом – интенсивно прогревается (Гидрометеорология …, 1990).
Ледовый покров толщиной до 1 м на севере образуется ежегодно, центральные и западные районы моря покрываются сплошным льдом только в суровые зимы. Средняя продолжительность ледового покрова 4–5 месяцев, в суровые зимы до 6 месяцев (Косарев, 1975).
Средняя соленость воды в Аральском море до 1961 г. была около 10.2 ‰ (Степанов, 1907; Бадер, 1934; Успенский, 1950; Блинов, 1956; Соловьева, 1959; Современный …, 1972; Бортник, 1977а, б; Гидрометеорология …, 1990). После 1961 г. началось быстрое увеличение солености, продолжающееся в Большом море и в настоящее время, причем рост солености вод моря имел тенденцию к ускорению. Так, за первые 10 лет (1961–1970) она увеличилась на 1.5 ‰, за вторые (1971–1980) – на 5.3 ‰, за третьи (1981–1990) – на 15.1 ‰. Возрастание солености обуславливается в основном уменьшением объема вод моря, так как солевая масса моря устойчива во времени.
Значительную роль в пространственном распределении солености в Аральском море для квазистационарного периода имел сток вод рек Амударьи (для южной и юго-западной части) и Сырдарьи (для северной части). Наименьшая соленость наблюдалась в приустьевых районах моря, максимальная – на восточном мелководье (рис. 3.3), где водные массы перемешивались слабо и преобладающее значение в формировании солености имело испарение (Блинов, 1956; Деньгина, 1959а; Ревина с соавт., 1970). В период нестационарного режима моря сокращение и, в отдельные годы, почти полное прекращение стока обусловило уменьшение, а затем и исчезновение относительно распресненных зон на устьевых взморьях рек Амударьи и Сырдарьи. Максимальные значения солености стали отмечаться в Малом море.
<![if !vml]>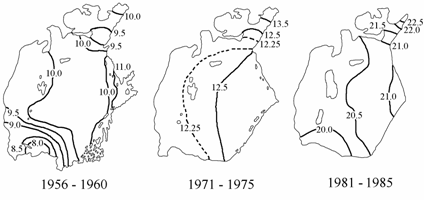 <![endif]>
<![endif]>
Рис. 3.3. Среднее многолетнее распределение солености на
поверхности Аральского моря летом (по: Гидрометеорология …, 1990), ‰.
В октябре 1989 г. в открытой части Аральского моря впервые была зарегистрирована соленость более 30 ‰, в сентябре 1990 г. соленость воды в Большом море уже на всей акватории превышала 30 ‰ (Цыцарин, 1991; Цыцарин, Бортник, 1991), а в августе 1992 г. в отдельных участках она превысила 40 ‰. Более интенсивное испарение на мелководьях привело к появлению хорошо выраженных зон максимальной солености у восточного побережья (40.0–40.5 ‰) и его северной части (41.0–41.2 ‰). В последнем районе существенное влияние на рост солености оказывает так же затрудненный водообмен с другими частями моря. Поверхностная соленость в августе 1992 г. в западной глубоководной котловине была в среднем на 3–4 ‰ меньше поверхностной солености в восточной части Большого моря из-за большего объема вод котловины и, следовательно, инерционности их. На юго-западную и южную часть моря оказывали распресняющее влияние воды р. Амударьи (рис. 3.4).
<![if !vml]>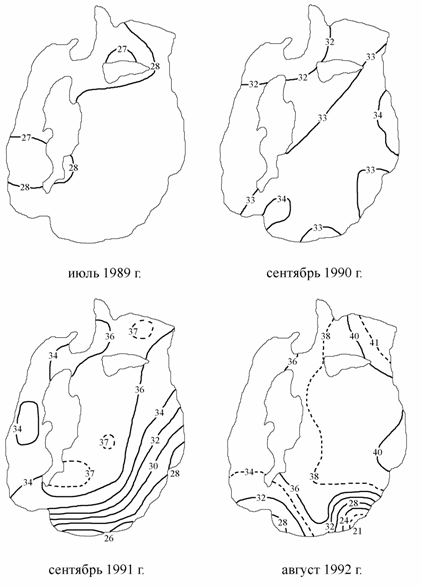 <![endif]>
<![endif]>
Рис. 3.4. Распределение солености на поверхности Большого моря (1989 г. по: Цыцарин, 1991; 1990 г. по: Цыцарин, Бортник, 1991; 1991–1992 гг. по: Цыцарин, Лобов, 1993б), ‰.
В Большом море в августе 1992 г. изменения солености по вертикали, даже в самом глубоком его районе – западной котловине, были невелики. Соленость там росла от поверхности до дна всего лишь на 0.2–0.8 ‰.
В Малом море в августе 1992 г. зона пониженной солености так же прослеживалась в районах, прилегающих к устью реки. Соленость воды, поступающей по р. Сырдарье, измеренная в 200 м выше впадения реки в море, оказалась равна 1.21 ‰. По сравнению с центральной частью Малого моря несколько повышенная соленость в поверхностном слое воды отмечалась в западной части. Наибольшая же соленость воды была в заливе Бутакова — 39–40 ‰ (рис. 3.5). В этом заливе, связанном с морем узким проливом, соленость с мая по август 1992 г. возросла на 3–4 ‰. Придонная вода в центральной части Малого моря в августе была более чем на 2 ‰ солонее вышележащих слоев. Существование слоя скачка солености, совпадающего по глубине залегания с термоклином, на такой небольшой глубине в конце теплого сезона объясняется пониженной ветровой активностью и значительным распреснением верхних слоев, приводящих к возрастанию плотностной устойчивости всей толщи воды.
Большое море в 1990-х годах значительно опережало в процессе осолонения Малое море. Так, средняя соленость вод Большого моря в августе 1992 г. составила 36.48 ‰, Малого моря – 28.65 ‰, разница 7.83 ‰.
Одновременно с осолонением моря происходило постоянное возрастание условной плотности морских вод, понижение температуры замерзания и снижение температуры наибольшей плотности воды.
<![if !vml]>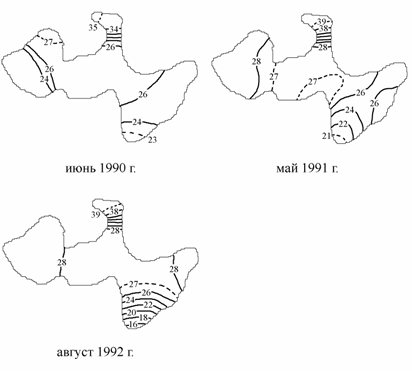 <![endif]>
<![endif]>
Рис. 3.5. Распределение солености на поверхности Малого моря (по: Лобов, 1994), ‰.
При квазистационарном режиме Аральского моря в нем, как солоноватом водоеме, процесс осенне-зимней конвекции протекал в две стадии. Поскольку для солоноватых вод температура наибольшей плотности выше температуры замерзания, то с началом осеннего выхолаживания вод начиналась первая стадия конвекции, которая продолжалась до достижения перемешанным слоем воды температуры наибольшей плотности. Затем, при дальнейшем охлаждении поверхностного слоя воды до температуры замерзания, плотность воды несколько уменьшалась, создавалась устойчивая стратификация и, несмотря на продолжавшуюся теплоотдачу, глубина конвенции не увеличивалась. При достижении температуры замерзания вод начиналась вторая стадия зимнего конвективного перемешивания, связанная с повышением плотности поверхностных вод в результате осолонения при льдообразовании (Косарев, 1975; Бортник, 1979).
После достижения солености воды в Аральском море 24.7 ‰ температура наибольшей плотности аральских вод стала равна температуре замерзания. Это привело к значительным изменениям характера протекания процесса осенне-зимнего конвективного перемешивания. Конвекция теперь протекает непрерывно в один этап до замерзания воды и появления льда. При этом вся масса вод выхолаживается до значительных отрицательных температур, равных температуре замерзания аральских вод соответствующей солености (Бортник, 1983; Гидрометеорология …, 1990).
При квазистационарном режиме Аральское море по солевому (ионному) составу воды занимало промежуточное положение между гидрокарбонатно-кальциевыми водами суши и хлоридно-натриевыми водами океана. Но все же вода Арала по соотношению солеобразующих ионов стояла ближе к типичным материковым водам (Блинов, 1956).
Процесс осолонения моря сопровождался метаморфизацией ионного состава за счет выпадения в осадок сульфата кальция, доломита и магнезиального кальцита (Квасов, Трофимец, 1976; Бортник, 1983). Анализ изменения солевого состава вод Аральского моря в разные периоды проводился неоднократно (Друмева, Цыцарин, 1984; Бортник, Буйневич, 1979, 1980; Бортник, Друмева, 1986, 1990; Гидрометеорология …, 1990; Цыцарин, Лобов, 1994, 1995, Лобов, 1994; и др.).
Ниже приведены изменения абсолютного (рис. 3.6) и относительного (рис. 3.7) содержания основных ионов, связь %-эквивалентных отношений основных ионов к хлору с суммой солей (рис. 3.8) и расчет концентраций гипотетических солей (рис. 3.9) в диапазоне солености 3–45‰. Для построения графиков был использован специально созданный массив данных о солевом составе вод Аральского моря (Цыцарин, Лобов, 1993а, 1994), обработанный с помощью специализированного пакета компьютерных программ, позволяющего производить расчеты в широком спектре изменений ионного состава (Лобов, Цыцарин, 1993).
<![if !vml]> <![endif]>
<![endif]>
<![if !mso]><![endif]>
<![if !vml]>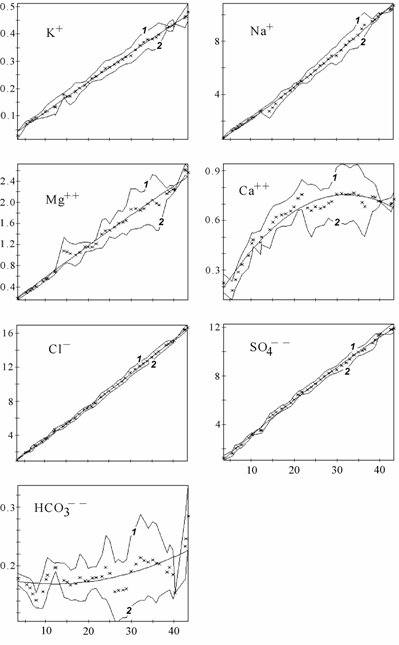 <![endif]>
<![endif]>
<![if !vml]> <![endif]>
<![endif]>
<![if !mso]><![endif]>
<![if !vml]>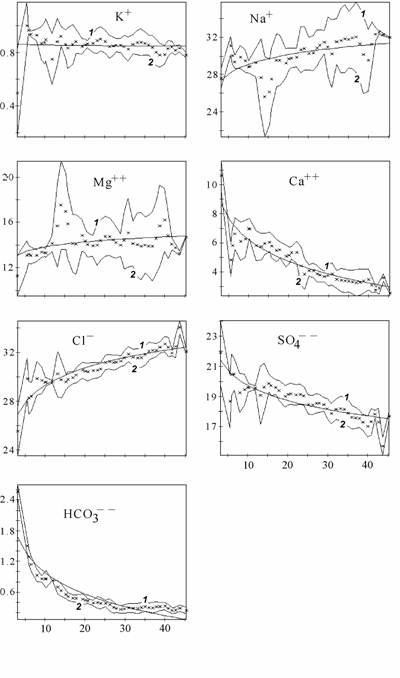 <![endif]>
<![endif]>
<![if !vml]>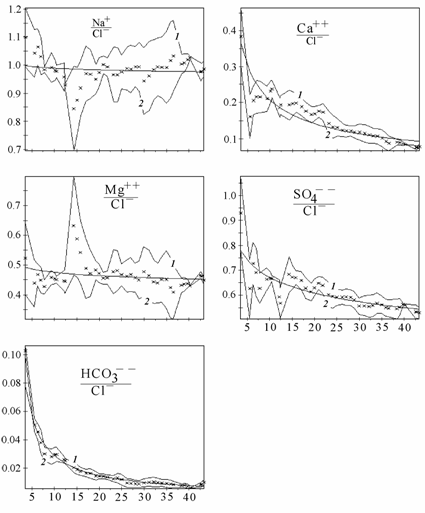 <![endif]>
<![endif]>
Рис. 3.8. Связь отношений относительных концентраций ионов к хлору (%-экв) с соленостью в воде Аральского моря. По оси абсцисс – соленость, ‰; по оси ординат – отношение ионов к хлору. Крестиком обозначено среднее значение в интервале. 1 и 2 – соответственно верхняя и нижняя границы интервалов ±d.
<![if !vml]> <![endif]>
<![endif]>
<![if !mso]><![endif]>
<![if !vml]>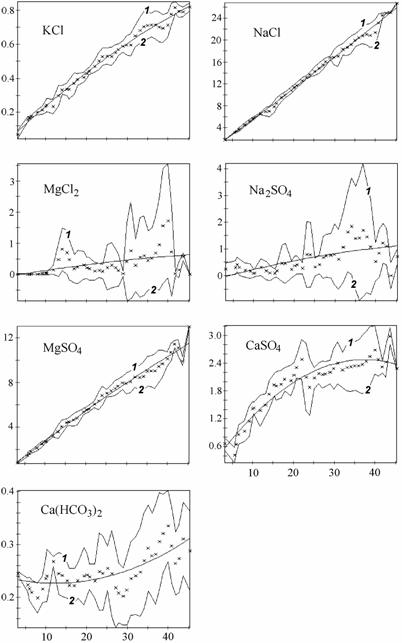 <![endif]>
<![endif]>
Увеличение солености воды Аральского моря привело солевую систему моря в состояние насыщения по отношению к некоторым солям. Солевая система потеряла устойчивость – теперь достаточно даже небольших изменений солености, температуры или pH воды, чтобы вызвать осаждение или растворение некоторого количества солей, при одновременных значительных изменениях в химическом составе воды (Цыцарин, Бортник, 1991).
В Аральском море одновременно с ростом общей солености и абсолютного содержания большинства ионов проходила седиментация карбонатов кальция и магния, гипса, которая определила понижение относительного содержания ионов кальция, магния, сульфатов, карбонатов и повышение содержания ионов хлора, натрия и калия, а также приводила к изменению отношения между хлорностью и соленостью (Лобов с соавт., 1995; см. рис. 3.6–3.9). На графиках (см. рис. 3.6–3.9) прослеживаются области изменения монотонности протекающих процессов, видимо соответствующие пороговым концентрациям, при которых наступает насыщение аральской воды очередной солью. При этом наблюдается резкое изменение равновесий в основных солевых системах, присутствующих в большинстве природных вод (Цыцарин, Лобов, 1994):
2NaCl + CaSO4 <==> CaCl2 + Na2SO4
2NaCl + MgSO4 <==> MgCl2 + Na2SO4
Ca(HCO3)2 <==> CaCO3 +H2O + CO2
Mg(HCO3)2 <==> MgCO3 +H2O + CO2
Установлены значимые уменьшения солезапасов Большого и Малого морей. Интегральные потери солей Аральского моря с 1978 по 1992 гг. составили за счет потери солей на осушающейся территории – 1.0 млрд. тонн и за счет выпадения в осадок непосредственно в море – 0.6 млрд. тонн (Цыцарин, 1991; Цыцарин, Бортник, 1991; Цыцарин, Лобов, 1994).
Ионный сток рек является основной составляющей приходной части солевого баланса Аральского моря (Гидрометеорология …, 1990). В 1961–1970 гг., несмотря на сокращение водного стока, суммарный ионный сток в Аральское море несколько увеличился и достиг 26.9 млн. т в год, главным образом за счет резкого, почти вдвое, возрастания общей минерализации вод р. Сырдарьи (табл. 3.1). В условиях дальнейшего сильного уменьшения водного стока суммарный ионный сток в Аральское море снизился в 1971–1980 гг. до 12.3, а в 1981–1985 гг. в среднем до 2.2 млн. т в год. В отдельные годы водный и ионный сток в море полностью отсутствовали. Минерализация вод Амударьи в 1981–1985 гг. в среднем была 1406, а Сырдарьи – 1844 мг/л, то есть по сравнению с периодом 1911–1960 гг. увеличилась соответственно 3.0 и 3.4 раза.
Таблица 3.1 Средняя многолетняя концентрация главных ионов, суммарная минерализация и ионный сток
рек Амударьи и Сырдарьи (по: Гидрометеорология …, 1990)
|
Период |
Единица измерения |
HCO3– |
SO42– |
Cl– |
Ca2+ |
Mg2+ |
Na++K+ |
Суммарная минерализация |
Ионный сток, |
|
Амударья |
|||||||||
|
1911–1960* |
мг/л |
153.5 |
104.9 |
74.1 |
83.1 |
11.2 |
43.7 |
470.8 |
17.74 |
|
|
%** |
18.53 |
16.08 |
15.39 |
35.92 |
7.98 |
6.10 |
100.00 |
|
|
1961–1970 |
мг/л |
124.8 |
131.0 |
85.0 |
62.0 |
19.0 |
61.0 |
482.8 |
16.93 |
|
|
% |
14.20 |
19.30 |
16.50 |
21.30 |
10.5 |
18.20 |
100.00 |
|
|
1971–1980 |
мг/л |
110.7 |
243.8 |
137.9 |
80.8 |
25.8 |
109.2 |
708.2 |
8.87 |
|
|
% |
8.34 |
23.62 |
18.04 |
18.45 |
9.79 |
21.76 |
100.00 |
|
|
1981–1985 |
мг/л |
145.3 |
619.3 |
357.4 |
127.8 |
74.2 |
315.2 |
1640.4 |
2.23 |
|
|
% |
4.69 |
29.43 |
19.88 |
12.58 |
12.03 |
25.39 |
100.00 |
|
|
Сырдарья |
|||||||||
|
1911–1960* |
мг/л |
186.1 |
164.4 |
40.3 |
87.6 |
20.6 |
43.8 |
542.8 |
6.05 |
|
|
% |
20.04 |
22.49 |
7.47 |
32.27 |
12.52 |
5.21 |
100.00 |
|
|
1961–1970 |
мг/л |
179.0 |
465.4 |
105.8 |
92.8 |
54.0 |
158.0 |
1055.0 |
9.96 |
|
|
% |
9.63 |
30.89 |
9.48 |
14.66 |
14.04 |
21.30 |
100.00 |
|
|
1971–1980 |
мг/л |
190.1 |
579.2 |
168.1 |
112.0 |
76.0 |
187.1 |
1312.5 |
3.59 |
|
|
% |
7.80 |
30.31 |
11.90 |
14.03 |
15.59 |
20.37 |
100.00 |
|
|
1981–1985 |
мг/л |
184.5 |
922.7 |
187.1 |
131.1 |
89.5 |
328.7 |
1843.6 |
– |
|
|
% |
5.49 |
34.92 |
9.59 |
11.88 |
13.37 |
24.75 |
100.00 |
|
Примечание: * – по: Блинов, 1956.
** – относительная форма молярной концентрации эквивалентов (% - экв).
Рост минерализации речных вод сопровождался резким изменением их ионного состава. Так, относительное содержание карбонатов в водах Амударьи и Сырдарьи снизилось в 4 раза, а их абсолютное содержание изменилось незначительно. Относительное уменьшение карбонатов в речном стоке компенсировалось увеличением абсолютного и относительного содержания хлоридов и значительным возрастанием абсолютного и относительного содержания сульфатов. Из катионов при небольшом росте абсолютного содержания кальция его относительное содержание уменьшилось втрое. При существенном росте абсолютного содержания магния его относительное содержание возросло незначительно. Наблюдался значительный рост абсолютного и относительного содержания щелочных металлов, в основном натрия. Таким образом, из гидрокарбонатно-кальциевых воды Амударьи и Сырдарьи, поступающие в море, трансформировались в сульфатно-натриевые. Возрастание суммарной минерализации речных вод и изменение их количественного состава связано с практически полным зарегулированием и хозяйственным использованием стока рек Амударьи и Сырдарьи, возрастанием в нем доли сильноминерализованных сбросных и дренажных вод (Гидрометеорология …, 1990).
Вода Аральского моря до зарегулирования стока рек была бедна основными биогенными элементами (Блинов, 1956), содержание фосфатов в среднем было 1–4 мкг/л при колебаниях от аналитического нуля до 23 мкг/л. Содержание нитритов было близко к нулю, средние концентрации нитратов не превышали 5 мкг/л, содержание аммонийного азота в поверхностном слое достигало 80 мкг/л. Среднее содержание кремнекислоты было 250 мкг/л, минимальные и максимальные концентрации – соответственно 120–800 мкг/л.
<![if !vml]>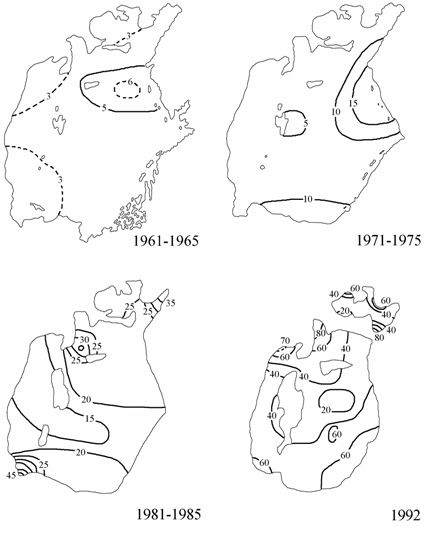 <![endif]>
<![endif]>
Рис. 3.10. Распределение фосфатов на поверхности Аральского моря (1961–1965, 1971–1975 и 1981–1985 гг. по: Гидрометеорология …, 1990; 1992 г. по: Лю Хун, 1993), мкг/л.
Данные о режиме фосфатов, нитритов и кремнекислоты Аральского моря за 1961–1985 гг. были обобщены И.В. Свиридовой (1990). Этот обзор и данные о содержании биогенных элементов в воде Аральского моря в августе 1992 г., были использованы при написании данного раздела.
Фосфаты. В летний период 1961–1965 гг. концентрации фосфатов на поверхности моря были близки к естественным — 1–6 мкг/л, максимальные концентрации наблюдались на северо-востоке Большого моря (рис. 3.10). К периоду 1971–1975 гг. произошло заметное увеличение концентраций фосфатов по сравнению с периодом 1961–1965 гг. На поверхности они изменялись уже от 5 до 18 мкг/л, зоны максимальных и повышенных концентраций (10–18 мкг/л) находились в восточной части Большого моря, на устьевых взморьях Амударьи и Сырдарьи. В последующие годы, особенно в 1981–1985 гг., произошло резкое увеличение средних концентраций фосфатов по всей акватории моря до 15–45 мкг/л.
В августе 1992 г. были отмечены наибольшие концентрации фосфатов за весь рассматриваемый период. Максимальные величины фосфатов отмечались в зоне влияния вод Амударьи и заливе Тщебас.
Нитриты. В 1961–1965 гг. в условиях близких к естественным в Аральском море летом средние концентрации нитритов изменялись в пределах 0.4–2.3 мкг/л (рис. 3.11). Минимальные концентрации нитритов были в центральном районе Большого моря, максимальные – в северо-западной части Большого моря.
В период 1966–1965 гг. отмечалось повсеместное заметное уменьшение содержания нитритов весной и летом.
В летнее время в период 1971–1975 гг. на большей части акватории содержание нитритов было ниже 1.0 мкг/л, опускаясь в отдельных районах до аналитического нуля. Максимальные концентрации нитритов (до 2.5 мкг/л) отмечались в северо-восточной части Большого
моря.
В 1976–1980 гг. содержание нитритов в летний период было выше, чем в 1971–1975 гг., а в весенний и осенний периоды нитриты отсутствовали в большинстве районов моря.
<![if !vml]>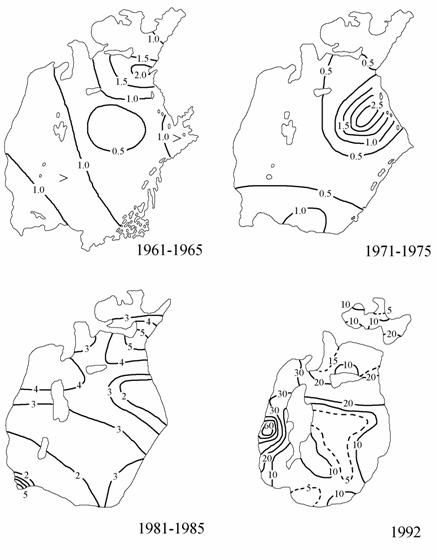 <![endif]>
<![endif]>
Рис. 3.11. Распределение нитритов на поверхности Аральского моря (1961–1965, 1971–1975 и 1981–1985 гг. по: Гидрометеорология …, 1990; 1992 г. по: Лю Хун, 1993), мкг/л.
В 1981–1985 гг. концентрации нитритов на поверхности значительно увеличились по сравнению с предшествующими пятилетками. В Малом море концентрации нитритов достигли 3–5 мкг/л.
Содержание нитритов на поверхности Аральского моря в августе 1992 г. изменялось от 5 до 64 мкг/л. Максимальные концентрации были приурочены к западной глубоководной впадине. В Малом море и на большей части акватории Большого моря содержание нитритов было в пределах 5–15 мкг/л (см. рис. 3.11).
Содержание нитратов и аммонийного азота на поверхности Аральского моря по сравнению с квазистационарным периодом также значительно увеличились. Распределение нитратов в августе 1992 г. было схожим с распределением нитритов, но при несколько больших величинах содержания. Максимум нитратов отмечен в западной глубоководной впадине, в районе южнее острова Барсакельмес их концентрации были 35–40 мкг/л, по восточной части Большого моря и большей части Малого моря – 10–20 мкг/л, на взморье Сырдарьи – 25 мкг/л (рис. 3.12А).
Максимум концентраций аммонийного азота был отмечен в районе мыса Изендыарал (160 мкг/л). Повышенное содержание аммонийного азота было характерно для западной глубоководной котловины (124 мкг/л), для зоны распреснения на юго-востоке Большого моря (60–75 мкг/л) и устьевого взморья Сырдарьи (140 мкг/л). В центральной и западной частях Малого моря содержание аммонийного азота было от 60 до 80 мкг/л (рис. 3.12Б).
Кремнекислота. В летнее время 1961–1965 гг. на поверхности концентрации кремнекислоты повышались до сезонного максимума во всех районах моря (рис. 3.13). Это было следствием влияния речного стока. Зоны повышенных концентраций (> 300 мкг/л) отмечались в восточной части Малого моря, на устьевом взморье Сырдарьи, в районе восточных мелководий, а также в юго-западной части Большого моря, где сказывалось влияние стока Амударьи.
В летнее время 1971–1975 гг. на поверхности Аральского моря по сравнению с тем же сезоном предшествующей пятилетки содержание кремнекислоты заметно возросло. Диапазон колебаний по акватории составлял 125–500 мкг/л. Максимальные концентрации находились на севере западного района, минимальные в западной части Малого моря.
<![if !vml]>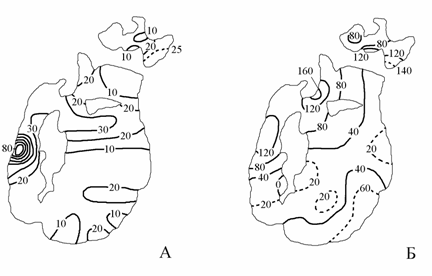 <![endif]>
<![endif]>
Рис. 3.12. Распределение нитратов (А) и аммонийного азота (Б) на поверхности Аральского моря в 1992 г. (по: Лю Хун, 1993), мкг/л.
В 1966–1970 гг. содержание и распределение кремнекислоты было близко к таковым в 1961–1965 гг.
В 1976–1980 гг. в ряде районов Аральского моря произошли изменения в режиме кремнекислоты, в основном, в сторону резкого увеличения ее концентрации.
В 1981–1985 гг. произошло дальнейшее увеличение концентрации кремнекислоты. Летом на поверхности содержание кремнекислоты в Малом море составляло 1100–1300 мкг/л, на устьевом взморье Сырдарьи – 800 мкг/л, в Аджибайском заливе – 1200 мкг/л. Минимальное (
В августе 1992 г. на поверхности Большого моря содержание кремнекислоты колебалось от 168 до 1665 мкг/л. Максимальные ее концентрации были приурочены к распресненной зоне у южного побережья (см. рис. 3.13). Повышенные значения кремнекислоты находились также на мелководьях южной части западной глубоководной котловины и на мелководьях южной части Большого моря. В заливе Тщебас повышенное содержание кремнекислоты (870 мкг/л) объясняется поступлением ее из скважины вместе с пресной водой. Пониженное содержание кремнекислоты отмечалось в водах центральной части Большого моря. В Малом море повышенным содержанием кремнекислоты отличались воды залива Бутакова и, особенно, устьевого взморья Сырдарьи. В западной и центральной частях Малого моря содержание кремнекислоты было относительно низкое – 200–400 мкг/л.
<![if !vml]>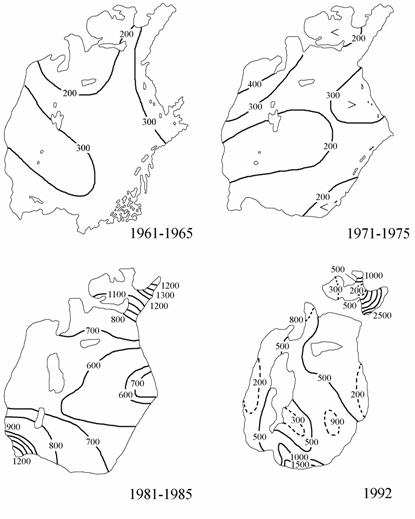 <![endif]>
<![endif]>
Рис. 3.13. Распределение кремния на поверхности Аральского моря (1961–1965, 1971–1975 и 1981–1985 гг. по: Гидрометеорология …, 1990; 1992 г. по: Лю Хун, 1993), мкг/л.
Основная составляющая приходной части баланса как растворенных, так и твердых форм биогенных веществ Аральского моря – биогенный сток Амударьи и Сырдарьи (Бруевич, Соловьева, 1957; Максимова с соавт., 1979; Бортник, 1980а).
В результате расширения удобряемых сельскохозяйственных угодий в бассейне Аральского моря в речных водах повысилось содержание растворенных биогенных веществ (табл. 3.2) и в первые годы (1961–1970) даже в условиях сокращения водного стока отмечалось заметное возрастание поступлений всех форм фосфора и азота, тогда как приток кремнекислоты остался на прежнем уровне (табл. 3.3). В 1971–1980 гг., несмотря на дальнейший рост концентраций большинства форм биогенных веществ в речном стоке, их приток, из-за прогрессирующего уменьшения водного стока, в Аральское море быстро сокращался. В 1981–1985 гг. в условиях незначительного водного стока приток биогенных веществ снизился до минимальных значений, а в отдельные годы водный сток и сток биогенных веществ в море полностью отсутствовал (Гидрометеорология …, 1990).
Так же как и для растворенных форм биогенных веществ, в 1961–1970 гг. при существенном сокращении водного стока отмечалось возрастание стока взвешенных форм биогенных веществ из-за возрастания мутности речных вод, главным образом Амударьи, и соответственно поступающего в море твердого стока (табл. 3.4). В 1977–1985 гг. при сохранении примерно тех же концентраций взвешенного вещества в речных водах из-за дальнейшего уменьшения водного стока поступление взвешенных форм биогенных веществ быстро достигло минимальных значений (Гидрометеорология …, 1990).
Таблица 3.2 Содержание растворенных биогенных веществ в речных водах (по табличным данным из: Гидрометеорология …, 1990), мг/м3
|
Период |
Водный сток, км3 |
P |
N |
Si |
|
Сырдарья |
||||
|
1911–1960 |
14.0 |
– |
1209–1279 |
4855–5300 |
|
1961–1965 |
9.6 |
200 |
2295 |
5000 |
|
1966–1970 |
10.6 |
99 |
1668 |
5200 |
|
1971–1975 |
5.3 |
128 |
2834 |
4200 |
|
1976–1980 |
0.3 |
86 |
2817 |
4400 |
|
Амударья |
||||
|
1911–1960 |
43.6 |
30–33 |
705–743 |
4756–4949 |
|
1961–1965 |
30.2 |
48 |
890 |
5800 |
|
1966–1970 |
36.6 |
48 |
1132 |
6000 |
|
1971–1975 |
16.2 |
27 |
702 |
4000 |
|
1976–1980 |
11.3 |
56 |
2268 |
4300 |
|
1981–1985 |
2.0 |
– |
2038 |
– |
Таблица 3.3 Среднее годовое поступление растворенных биогенных веществ в Аральское море (по табличным данным из: Гидрометеорология …, 1990), тыс. т
|
Период |
Речной сток |
Осадки |
||||||||||
|
P–PO4 |
Pорг |
Pвал |
N–NO2 |
N–NO3 |
N–NH4 |
Nмин |
Nорг |
Nвал |
Si |
N–NO3 |
N–NH4 |
|
|
1911–1960 |
0.5 |
1.0 |
1.5 |
– |
20.9 |
3.1 |
24.0 |
24.7 |
48.7 |
288.0 |
1.8 |
1.8 |
|
1961–1965 |
1.3 |
1.9 |
3.2 |
0.2 |
12.6 |
3.1 |
15.9 |
31.1 |
47.0 |
231.2 |
1.5 |
5.6 |
|
1966–1970 |
1.2 |
1.4 |
2.6 |
0.3 |
20.0 |
15.8 |
36.1 |
23.3 |
59.4 |
296.2 |
1.6 |
4.0 |
|
1971–1975 |
0.5 |
0.5 |
1.0 |
0.2 |
8.2 |
7.0 |
15.4 |
8.6 |
24.0 |
115.9 |
2.9 |
3.1 |
|
1976–1980 |
0.3 |
0.4 |
0.7 |
0.3 |
17.7 |
3.6 |
21.6 |
6.6 |
28.2 |
48.1 |
1.9 |
4.8 |
|
1981–1985 |
– |
– |
– |
<0.1 |
2.2 |
0.2 |
2.4 |
1.6 |
4.0 |
– |
2.0 |
3.4 |
Высокое абсолютное и относительное содержание кислорода в воде Аральского моря на всех глубинах в течение всего года отмечалось рядом авторов (Бенинг, 1935; Блинов, 1956; Соловьева, 1959; Косарев, 1975; Бортник, Чудаков, 1978), при этом особенностью Арала являлось пересыщение кислородом придонных слоев, за исключением района глубоководной впадины, где на глубинах более 40 м относительное содержание кислорода было чуть меньше 100 %. По мере осолонения моря относительное содержание кислорода в придонных слоях воды глубоководной впадины с обычных 90 % понизилось до 60 % в 1981–1985 гг. (Гидрометеорология …, 1990). В августе 1992 г., по данным ГОИНа, в придонных слоях глубоководной котловины вновь наблюдалось 90 % относительное содержание кислорода.
Таблица 3.4 Среднее годовое поступление твердых биогенных веществ в Аральское море (по табличным данным из: Гидрометеорология …, 1990), тыс. т
|
Период |
Речной сток |
Атмосферная пыль |
|||
|
|
P |
N |
Si |
P |
N |
|
1911–1960 |
56.0 |
82.0 |
29244 |
1.7 |
5.0 |
|
1961–1965 |
61.4 |
59.3 |
24287 |
1.6 |
4.7 |
|
1966–1970 |
115.5 |
112.1 |
45585 |
1.5 |
4.4 |
|
1971–1975 |
37.7 |
36.5 |
14914 |
1.4 |
4.2 |
|
1976–1980 |
14.5 |
14.3 |
5750 |
2.2 |
6.5 |
|
1981–1985 |
1.5 |
1.4 |
588 |
2.0 |
5.7 |
Для всей акватории Аральского моря характерна щелочная реакция среды. До 1961 г. среднее значение pH в зависимости от сезона колебалось от 8.2 до 8.4, достигая на мелководных с повышенной соленостью участках восточного побережья 9 (Блинов, 1956; Соловьева, 1959). В августе 1992 г. средняя величина pH была 8.6 (рис. 3.14).
При квазистационарном режиме в Аральском море, в отличие от других внутренних морей, органическое вещество было в основном аллохтонного происхождения (Максимова, 1973), 53.5 % от общего годового прихода поступало со стоком рек, 42.8 % – продуцировали макрофиты и 3.7 % – фитопланктон. Около 90 % приносимого реками органического вещества поступало во взвешенном состоянии, большая часть его выпадала вблизи устьев рек и выключалось из общего круговорота органического вещества. 62.3 % от общего годового прихода органического вещества оседало в грунт и 36.7 % минерализовалось.
Донные осадки (грунты) Аральского моря формировались за счет взвешенных наносов, выносимых реками, эоловых наносов, а также в результате отмирания и минерализации организмов планктона и бентоса.
До изъятия стока рек в донные отложения моря ежегодно переходило около 130 млн. т осадочных веществ, терригенная часть составляла 85.4 %, хемогенные и биогенные продукты 10.4 и 4.2 % соответственно (Резников, 1976; Хрусталев с соавт., 1977). Причем основу седиментационного материала составляли выносы рек Амударьи и Сырдарьи (72.3 %), а абразия берегов и эоловые наносы имели меньшее значение.
<![if !vml]> <![endif]>
<![endif]>
Рис. 3.14. Распределение pH на поверхности моря летом (1961–1965 гг. по: Гидрометеорология …, 1990; 1992 г. по: Лю Хун, 1993).
На основании гранулометрического, минералогического состава и условий образования была составлена схема (рис. 3.15) донных осадков Аральского моря (Хрусталев с соавт., 1977). Пески были приурочены в основном к береговой зоне Аральского моря, тем областям, где происходило постоянное взмучивание донных осадков. Оолитовые известковые пески протягивались от банки Бенинга на юг до острова Лазарева, их распространение ограничивалось 20-ти метровой изобатой. Алевритовые илы встречались на восточном мелководье, на различных глубинах придельтовых областей, в открытой части моря, северных заливах и Малом море, располагаясь в зонах, где воздействие волнения на дно крайне ослаблено. Глинистые илы были расположены в придельтовых областях, северных заливах, в открытой части водоема и Малом море. В придельтовых районах зоны распространения глинистых илов были крайне неустойчивы и зависели от сезонности и гидродинамической обстановки. Глинисто-известковые илы были приурочены к западной и центральной глубоководным впадинам, представляли собой полужидкую массу темно-серого или черного цветов часто с запахом сероводорода.
<![if !vml]> <![endif]>
<![endif]>
Рис. 3.15. Схема донных осадков Аральского моря (по: Хрусталев с соавт., 1977).
1 - пески; 2 – оолитовые известковые пески; 3 – крупные алевриты; 4 –мелкоалевритовые илы; 5 – глинисто-известковые илы; 6 – глинистые илы дельт, северных заливов и Малого моря.
В практической работе мы оценивали грунты на основании механического состава и по цвету (рис. 3.16). Илистый песок и песчанистый ил соответствовали крупным алевритам и мелкоалевритовым илам. Цвет илов Аральского моря зависит, в основном, от окислительно-восстановительных процессов, протекающих в грунте. При высыхании на воздухе черный или темно-серый илы светлеют и по цвету почти не отличаются от высушенного серого ила. Появление коричнево-серых илов обуславливалось седиментацией осадков, влекомых реками; площади их изменчивы и целиком и полностью зависят от объема стока.
<![if !vml]> <![endif]>
<![endif]>
Рис. 3.16. Схема распределения грунтов в Аральском море.
1 – песок, 2 – илистый песок, 3 – песчанистый ил, 4 – коричневый и коричнево-серый илы, 5 – серый ил, 6 – темно-серый ил, 7 – черный ил.
По мере обмеления Аральского моря в центральной глубоководной котловине перестали встречаться темно-серые илы, имевшие уже в 1978 г. ограниченное распространение. Мелководные участки по восточному побережью и возле гряды островов, занятые песками и илистыми песками, осушились. Преобладающими грунтами стали серые илы, резко сократились площади занятые черными и темно-серыми илами. Следует отметить, что при возобновлении исследований в 1989 г. мы на большей части дна Большого моря обнаружили корку соли (1–3 мм), сцементировавшую поверхность грунтов. В ряде случаев корка соли даже препятствовала работе дночерпателя Петерсена, не давая ему погрузиться в грунт. В последующие годы солевая корка как бы ослабла, поверх нее произошла седиментация взвешенных частиц и затруднений с взятием грунта не было.
Регулярные наблюдения за состоянием химического загрязнения вод Аральского моря по сети станций Общегосударственной системы наблюдений Госкомгидромета СССР (см. Бортник, Гейдарова, 1991) проводятся с 1970 г., с перерывом в 1987–1988 гг.
Как отмечают В.Н. Бортник и Н.С. Гейдарова (1991), в начале 1970?х годов к основным источникам загрязнения вод моря относились речные воды Амударьи и Сырдарьи, рыболовный и транспортный флот, Аральский морской порт, Аральский и Муйнакский рыбокомбинаты. С середины 1970-х к началу 1980-х годов, одновременно с резким сокращением стока рек, были введены очистные сооружения на Муйнакском и Аральском рыбокомбинатах, прекратились транспортные перевозки и рыбный промысел, закрылся Аральский морской порт. В 1980-х годах загрязняющие вещества поступали в Аральское море только от небольшого количества оставшихся судов, нерегулярно с речным стоком Сырдарьи и Амударьи, а также с небольшим количеством коллекторно-дренажных вод, доходящих до моря, и при аварийных сбросах вследствие прорыва земляных дамб в нижнем течении Амударьи и Сырдарьи.
Существенное уменьшение поступления в море загрязняющих веществ привело к снижению уровня его загрязнения. Так, по средним показателям загрязняющих веществ (нефтяные углеводороды, фенолы, синтетические поверхностно активные вещества, хлорорганические пестициды, тяжелые металлы) в 1970–1979 гг. воды Аральского моря классифицировались как "чрезвычайно грязные", в 1980–1986 гг. – как "условно чистые" и в 1989 г. – как "загрязненные". Очевидно, увеличение загрязнения Аральского моря в 1989 г. было обусловлено относительно многоводным 1988 г. (Бортник, Гейдарова, 1991). В начале 1990-х годов Аральское море отличалось меньшим уровнем пестицидного загрязнения по сравнению с многими крупными рыбохозяйственными водоемами Казахстана (Амиргалиев с соавт., 1993).
Таким образом, произошли существенные изменения Аральского моря как среды обитания гидробионтов по сравнению с периодом его квазистационарного режима. Помимо изменения морфометрических характеристик, резко повысилась соленость воды, уменьшилось поступление биогенных элементов, понизилась температура воды в зимний период. Претерпели изменения и воды впадающих рек, из гидрокарбонатно-кальциевых они трансформировались в сульфатно-натриевые, в них возросло содержание биогенных и загрязняющих веществ. По существу Аральское море в современном его состоянии – совершенно иная среда обитания по сравнению с периодом его квазистационарного режима, а в недалеком будущем возможен переход Большого моря в гипергалинный водоем.
Той флоры и той фауны, которые описывались исследователями первой половины XX века (Берг, 1908; Бенинг, 1934, 1935; Никольский, 1940; Яблонская, 1960; Доброхотова, 1971; Пичкилы, 1971, Атлас …, 1974) как типично аральские, в Аральском море больше нет. Все генеративно-пресноводные и солоноватоводные виды водных растений, беспозвоночных и рыб вымерли, а их место занято небольшим числом интродуцированных эвригалинных морских видов, либо малочисленных ранее аборигенных эвригалинных и галофильных видов, либо вакантно.
Так, в составе фитопланктона в 1995 г. насчитывалось 243 вида (Orlova et al., 1998) против 306, зарегистрированных Л.О. Пичкилы (1971), причем резко изменилось соотношение между отдельными группами водорослей. Из 37 видов макрофитов (Доброхотова, 1971) к середине 1990-х годов сохранилось 5 видов (Orlova et al., 1998): Zostera noltei Hornem, Ruppia cirrhosa (Petagna) Grande, Chaetomorpha linum (Mull.), Cladophora glomerata (L.) Kutz., Cl. fracta (Mull. ex. Vahl.) Kutz.
В составе зоопланктона на 1992 г. сохранилось 9 видов. Обеднение видового состава зоопланктона Аральского моря в результате осолонения воды в основном произошло при солености 12–14 и 25–28 ‰. Из всего разнообразия видов сохранились: среди коловраток – Synchaeta vorax Rouss.; среди ветвистоусых: в более пресном Малом море – Podonevadne camptonyx (G. Sars), в более соленом Большом море – Moina mongolica Daday; среди веслоногих ракообразных – Calanipeda aquaedulcis Kritsch., Halicyclops rotundipes aralensis Borutzky, Schizopera aralensis Borutzky, Nitocra lacustris (Schmank.); а также личинки двустворчатых моллюсков – Syndosmya и Cerastoderma. Основу численности и биомассы зоопланктонного сообщества формируют акклиматизанты: Calanipeda aquaedulcis и личинки Syndosmya (Андреев, 1999).
В составе ихтиофауны за все годы наблюдений было встречено 33 вида рыб, из которых 13 акклиматизанты. К 1994 г. сохранилось 9 видов: Clupea harengus membras Linné, Atherina boyeri caspia (Eichw.), Pungitius platygaster aralensis (Kessler), Pomatoschistus caucasicus (Kawrajsky), Neogobius melanostomus affinis (Eichw.), Neogobius kessleri gorlap Iljin, Neogobius fluviatilis pallasi (Berg), Proterorhinus marmoratus (Pallas), Platichthys flesus luscus (Pallas), из которых 8 акклиматизанты, причем Neogobius kessleri gorlap и Proterorhinus marmoratus в последние годы в уловах не отмечались (Андреев, 1999).
Изменения видового состава и количественного развития зообентоса следует рассмотреть более подробно (см. также Андреев, Андреева, 1997а), так как именно на фоне дестабилизации биотического окружения, особенно на дне моря, и произошли события, рассматриваемые в данной работе.
При нормальной солености воды открытого моря в составе зообентоса Арала с 1874 по 1971 гг. было отмечено 7 видов кольчатых червей, 5 – высших ракообразных, 17 – насекомых и 12 видов и подвидов моллюсков (табл. 3.5).
Единственный представитель многощетинковых червей – средиземноморско-атлантический вид Nereis diversicolor акклиматизирован в Аральском море в 1960–1961 гг., отмечается в море с 1963 г. (Кортунова, 1968, 1970).
Все малощетинковые черви, встреченные в Аральском море, согласно О.В. Чекановской (1962), широко распространенные виды, свойственные прежде всего пресным водам разного типа, но выдерживающим ту или иную концентрацию солей, некоторые вплоть до океанической солености.
Таблица 3.5 Многолетние изменения видового состава зообентоса открытой части Аральского моря (по: Андреев, Андреева, 1997а)
|
Вид |
1971 |
1976–1977 |
1980 |
1989–1992 |
|
Nereis diversicolor O.F. Müller |
+ |
+ |
+ |
+ |
|
Nais elinguis Müller |
+ |
– |
– |
– |
|
Paranais simplex Hrabe |
+ |
– |
– |
– |
|
Limnodrilus helveticus Piguet |
+ |
– |
– |
– |
|
Potamothrix bavaricus (Oeschm.) |
+ |
– |
– |
– |
|
Psammoryctides albicola (Mich.) |
+ |
– |
– |
– |
|
Lumbricilus lineatus (Müller) |
+ |
– |
– |
– |
|
Paramysis intermedia (Czern.) |
+ |
+ |
– |
– |
|
P. lacustris (Czern.) |
+ |
+ |
– |
– |
|
Dikerogammarus aralensis (Uljan.) |
+ |
– |
– |
– |
|
Palaemon elegans Rathke |
+ |
+ |
+ |
+ |
|
Rhithropanopeus harrisii tridentatus (Maitl.) |
+ |
+ |
+ |
+ |
|
Agrypnetes crassicornis McL. |
+ |
– |
– |
– |
|
Oecetis intima McL. |
+ |
+ |
– |
– |
|
Pelopia villipennis Kieff. |
+ |
– |
– |
– |
|
Procladius ferrugineus Kieff. |
+ |
– |
– |
– |
|
Cricotopus gr. silvestris F. |
+ |
– |
– |
– |
|
Tanytarsus gr. lobatifrons Kieff. |
+ |
– |
– |
– |
|
T. gr. gregarius Kieff. |
+ |
– |
– |
– |
|
T. gr. lauterborni Kieff. |
+ |
– |
– |
– |
|
T. gr. exiguus Joh. |
+ |
– |
– |
– |
|
Polypedilum gr. scalaenum Schr. |
+ |
– |
– |
– |
|
Cryptochironomus supplicans Meig. |
+ |
– |
– |
– |
|
Cr. gr. defectus Kieff. |
+ |
– |
– |
– |
|
Cr. gr. viridulus F. |
+ |
– |
– |
– |
|
Cr. gr. conjugens Kieff. |
+ |
– |
– |
– |
|
Chironomus behningi Goetgh. |
+ |
– |
– |
– |
|
Ch. halophilus Kieff. |
+ |
+ |
+ |
– |
|
Ch. salinarius Kieff. |
+ |
+ |
+ |
– |
|
Dreissena obtusecarinata (Andr.) |
+ |
+ |
– |
– |
|
D. caspia pallasi (Andr.) |
+ |
+ |
+ |
– |
|
Cerastoderma rhomboides rhomboides (Lam.) |
+ |
? |
– |
– |
|
C. isthmicum Issel |
+ |
+ |
+ |
+ |
|
Hypanis vitrea bergi Star. |
+ |
+ |
– |
– |
|
H. sidorovi Star. |
+ |
+ |
– |
– |
|
H. minima minima (Ostr.) |
+ |
+ |
– |
– |
|
Syndosmya segmentum Reclus |
+ |
+ |
+ |
+ |
|
Theodoxus pallasi Ldh. |
+ |
+ |
+ |
– |
|
Caspiohydrobia husainovae Star. |
+ |
+ |
+ |
+ |
|
C. conica (Logv. et Star.) |
+ |
+ |
+ |
+ |
|
C. obrutchevi Star. et Andreeva |
+ |
+ |
+ |
+ |
|
C. convexa (Logv. et Star.) |
? |
+ |
+ |
+ |
|
C. kazakhstanica Star. et Andreeva |
? |
+ |
+ |
+ |
|
C. aralensis Star. et Andreeva |
? |
+ |
+ |
+ |
|
C. nikitinskii Star. et Andreeva |
? |
+ |
+ |
+ |
|
C. parva (Logv. et Star.) |
? |
+ |
+ |
+ |
|
C. dubia (Logv. et Star.) |
? |
+ |
+ |
+ |
|
C. curta (Logv. et Star.) |
? |
+ |
+ |
+ |
|
C. gemmata (Kol.) |
? |
+ |
+ |
+ |
|
C. nikolskii Star. et Andreeva |
? |
+ |
+ |
– |
|
C. bergi Star. et Andreeva |
? |
+ |
+ |
+ |
|
C. oviformis (Logv. et Star.) |
? |
+ |
+ |
+ |
|
C. subconvexa (Logv. et Star.) |
? |
+ |
+ |
+ |
|
C. grimmi (Cless. et W. Dyb.) |
? |
+ |
+ |
+ |
|
C. chrysopsis (Kol.) |
? |
+ |
+ |
+ |
|
C. cylindrica (Logv. et Star.) |
? |
+ |
+ |
+ |
|
C. behningi Star. et Andreeva |
? |
+ |
+ |
+ |
|
C. sidorovi Star. et Andreeva |
? |
+ |
+ |
+ |
|
C. pavlovskii Star. et Izzat. |
? |
+ |
+ |
+ |
|
C. tadzikistanica Star. et Izzat. |
? |
+ |
+ |
+ |
|
C. sogdiana Star. et Izzat. |
? |
+ |
+ |
+ |
Примечание: список видов 1971 г. составлен по: «Атлас ...» (1974); Ch. salinarius приведен Р.С. Деньгиной (1959); C. obrutchevi описан по сборам 1956 г. (Старобогатов, Андреева, 1981); 1976–1977 и 1980 гг. по: Андреева (1989).
Из высших ракообразных только Dikerogammarus aralensis был аборигенным видом. Он, помимо Арала и озер низовьев впадающих в него рек, встречается в Каспии (Романова, 1974). Остальные виды были акклиматизированы: каспийские мизиды Paramysis intermedia и P. lacustris в 1958–1960 гг., отмечаются с 1961 г. (Галактионова, Бинтинг, 1964); европейско-африканская креветка Palaemon elegans завезена при интродукции кефали, впервые обнаружена в 1957 г. (Коновалов, 1959); североатлантический краб Rhithropanopeus harrisii tridentatus завезен при акклиматизации Calanipeda aquaedulcis (Мордухай-Болтовской, 1972), встречен в навигацию 1976 г. (Андреев, Андреева, 1988).
Ручейники в Аральском море были представлены видами, широко распространенными в солоноватых водоемах Европейской части России, Казахстана, Средней Азии и Монголии (Сибирцева, 1974).
Личинки хирономид были представлены в Арале видами пресноводных родов, из которых виды родов Cricotopus и Chironomus могут встречаться при большом осолонении (Панкратова, 1970).
Среди аральских моллюсков выделяют (Мордухай-Болтовской, 1960, 1978; Яблонская, 1974) группу «каспийских» видов (все Dreissena, Hypanis и Theodoxus pallasi) и группу средиземноморско-атлантических видов (Cerastoderma и Syndosmya segmentum). Последний вид был вселен в Арал в 1960–1961 и 1963 гг., отмечается с 1967 г. (Кортунова, 1970). Моллюсков рода Caspiohydrobia, относившихся ранее к автохтонам Каспия (Мордухай-Болтовской, 1960), теперь считают сформировавшимися в осолоненных водоемах внутренних частей Азии, откуда шло их расселение в Арал и Каспий (Старобогатов, Андреева, 1981; Андреева, 1987).
Таким образом, к моменту резкого падения уровня моря и повышения солености воды, зообентос Аральского моря был представлен небольшим числом наиболее эвригалинных видов пресноводного происхождения, «каспийскими» и средиземноморско-атлантическими видами и видами, сформировавшимися во внутренних частях Азии. Следует подчеркнуть, что массового развития в бентосе достигали пресноводные виды (Chironomidae) и «каспийские» моллюски родов Dreissena и Hypanis (Никитинский, 1933; Хусаинова, 1954; Яблонская, 1960).
В период начального осолонения Аральского моря (1972–1981 гг.) сокращение стока рек и повышение солености воды неблагоприятно сказалось на пресноводных и солоноватоводных «каспийских» видах, началось их быстрое выпадение из фауны (Андреев с соавт., 1978; Андреева, 1978, 1983; Андреев, Андреева, 1979; Губа, Андреев, Андреева с соавт., 1981). Если по списку видов по состоянию на 1971 г. (см. табл. 3.5) в зообентосе открытого моря при его нормальной солености отмечался 41 вид и подвид, то к 1976–1977 гг., из этого списка сохранилось 20, а к 1980 г. осталось 12 видов. Кроме того, по сборам 1976–1980 гг. для фауны Арала было отмечено еще 20 видов моллюсков рода Caspiohydrobia (Старобогатов, Андреева, 1981; Андреева, 1987). Нахождение большого числа видов в казалось бы хорошо изученном водоеме связано с расселением моллюсков из удаленных осолоненных заливов, остававшихся слабо исследованными в фаунистическом отношении, в связи с возникновением благоприятных условий обитания при повышении солености воды в море (Андреева, 1984).
В начальный период прогрессирующего осолонения (1982–1988 гг.) из фауны Аральского моря выпали Dreissena caspia pallasi, Theodoxus pallasi, Chironomus halophilus, Ch. salinarius, Caspiohydrobia nikolskii. По сборам зообентоса в Малом море первым исчез Dreissena caspia pallasi, затем остальные виды; впрочем, 2 экземпляра длиной около 5 мм ближе не определенных Chironomidae были отловлены в августе 1989 г. в Малом море (Андреев, Андреева, Филиппов, 1990), после чего они перестали встречаться как в Малом, так и в Большом море.
В 1989–1994 гг. в Малом море, помимо упоминавшихся Chironomidae, были встречены только Nereis diversicolor, Syndosmya segmentum, Cerastoderma ? isthmicum, Palaemon elegans и 18 видов Caspiohydrobia: C. behningi, C. oviformis, C. kazakhstanica, C. husainovae, C. tadzikistanica, C. aralensis, C. obrutchevi, C. dubia, C. nikitinskii, C. convexa, C. cylindrica, C. chrysopsis, C. sidorovi, C. pavlovskii, C. sogdiana, C. gemmata, C. parva, C. grimmi; в Большом море в 1989–1992 гг. к ним добавились Rhithropanopeus harrisii tridentatus, Caspiohydrobia bergi, C. subconvexa, C. curta, C. conica (Andreev et al., 1992; Андреев, Андреева, 1996, 1997а).
Как показал проведенный нами (Андреев, Андреева, 1997а) анализ, обеднение видового состава зообентоса Аральского моря в результате осолонения воды (для Dikerogammarus aralensis, возможно, и вследствие конкуренции с Palaemon elegans) в основном произошло при солености 12–14 и 25–28 ‰. В составе зообентоса остались только эвригалинные виды морского происхождения: Nereis diversicolor, Syndosmya segmentum, Cerastoderma ? isthmicum, Palaemon elegans, Rhithropanopeus harrisii tridentatus и галофильные виды материковых вод – представители рода Caspiohydrobia.
Рассматривая все имеющиеся сведения о биомассе зообентоса Арала (табл. 3.6), нетрудно убедиться, что до середины 1960-х годов наблюдалось относительное постоянство как общей биомассы зообентоса, так и биомассы отдельных групп организмов. Небольшие флюктуационные колебания биомассы зообентоса имели достоверную коррелятивную связь с речным стоком, причем при увеличении выноса взвешенных веществ в море происходило увеличение биомассы пелофильных личинок хирономид и уменьшение биомассы видов (Dreissena, Dikerogammarus aralensis), предпочитающих более плотные грунты (Никольский, Фортунатов, 1950; Яблонская, 1960).
Таблица 3.6 Средняя биомасса зообентоса открытой части Аральского моря летом (по: Андреев, Андреева, 1997а), г/м2
|
Годы |
Nereis |
Dikerogammarus |
Chironomidae |
Dreissena |
Hypanis |
Cerastoderma |
Syndosmya |
Theodoxus |
Caspiohydrobia |
Прочие |
Всего |
|
1930 |
– |
0.5 |
4.2 |
12.1 |
5.5 |
0.3 |
– |
– |
– |
0.3 |
22.9 |
|
1932 |
– |
0.1 |
2.6 |
7.8 |
4.6 |
0.5 |
– |
0.1 |
0.4 |
0.8 |
16.9 |
|
1933 |
– |
0.6 |
3.8 |
7.2 |
6.6 |
0.4 |
– |
0.2 |
<0.1 |
0.4 |
19.6 |
|
1936 |
– |
0.4 |
5.3 |
8.3 |
7.0 |
– |
– |
0.2 |
1.2 |
0.6 |
23.0 |
|
1937 |
– |
0.1 |
5.7 |
7.4 |
6.3 |
– |
– |
0.2 |
1.2 |
1.0 |
21.0 |
|
1938 |
– |
0.2 |
5.4 |
6.3 |
6.3 |
– |
– |
0.1 |
1.4 |
0.3 |
20.0 |
|
1939 |
– |
0.2 |
4.6 |
5.4 |
5.4 |
– |
– |
0.1 |
1.2 |
0.1 |
17.0 |
|
1940 |
– |
0.1 |
3.3 |
3.8 |
3.8 |
– |
– |
0.1 |
0.9 |
<0.1 |
12.0 |
|
1946 |
– |
x |
x |
x |
x |
x |
– |
x |
x |
x |
24.4 |
|
1948 |
– |
x |
x |
x |
x |
x |
– |
x |
x |
x |
19.4 |
|
1954 |
– |
0.2 |
8.6 |
8.4 |
8.2 |
<0.1 |
– |
0.1 |
<0.1 |
0.3 |
25.8 |
|
1956 |
– |
0.1 |
10,5 |
12.0 |
5.3 |
0.5 |
– |
1.4 |
0.1 |
<0.1 |
29.9 |
|
1957 |
– |
<0.1 |
6.3 |
3.6 |
3.9 |
1.6 |
– |
<0.1 |
0.1 |
<0.1 |
15.6 |
|
1959 |
– |
x |
x |
x |
x |
x |
– |
x |
x |
x |
23.1 |
|
1960 |
– |
0.1 |
9.7 |
x |
x |
x |
– |
x |
x |
x |
27.9 |
|
1961 |
– |
0.1 |
8.1 |
8.0 |
7.6 |
1.0 |
– |
1.0 |
0.8 |
0.1 |
26.7 |
|
1962 |
– |
0.2 |
12.2 |
14.9 |
7.1 |
3.2 |
– |
0.4 |
2.4 |
0.2 |
40.6 |
|
1963 |
– |
0.3 |
5.4 |
12.6 |
5.1 |
3.0 |
– |
0.2 |
2.5 |
0.2 |
29.3 |
Окончание табл. 3.6
|
Годы |
Nereis |
Dikerogammarus |
Chironomidae |
Dreissena |
Hypanis |
Cerastoderma |
Syndomya |
Theodoxus |
Caspiohydrobia |
Прочие |
Всего |
|
1964 |
<0.1 |
0.1 |
4.7 |
6.9 |
1.5 |
2.4 |
– |
0.2 |
2.0 |
0.1 |
17.9 |
|
1965 |
0.2 |
0.1 |
3.4 |
6.1 |
1.4 |
2.2 |
– |
0.5 |
1.9 |
0.2 |
16.0 |
|
1966 |
0.6 |
<0.1 |
1.4 |
3.1 |
1.5 |
0.8 |
– |
0.2 |
1.7 |
<0.1 |
9.3 |
|
1967 |
1.1 |
<0.1 |
0.9 |
2.3 |
0.2 |
2.1 |
0.1 |
0.1 |
3.1 |
<0.1 |
9.9 |
|
1968 |
2.8 |
<0.1 |
0.6 |
2.8 |
0.9 |
3.9 |
1.1 |
0.1 |
2.2 |
<0.1 |
14.4 |
|
1969 |
2.3 |
<0.1 |
0.5 |
1.0 |
0.8 |
4.3 |
2.1 |
<0.1 |
0.7 |
<0.1 |
11.7 |
|
1970 |
5.9 |
<0.1 |
0.4 |
1.0 |
0.4 |
3.4 |
12.1 |
0.1 |
1.0 |
<0.1 |
24.3 |
|
1971 |
6.5 |
<0.1 |
0.3 |
0.9 |
1.3 |
3.1 |
11.9 |
0.1 |
1.6 |
<0.1 |
25.7 |
|
1972 |
9.7 |
<0.1 |
0.1 |
1.4 |
2.0 |
7.1 |
11.3 |
0.1 |
1.4 |
<0.1 |
33.1 |
|
1973 |
7.4 |
– |
<0.1 |
0.1 |
0.6 |
7.2 |
15.2 |
<0.1 |
0.1 |
– |
30.6 |
|
1974 |
10.3 |
– |
– |
0.9 |
0.5 |
10.4 |
30.4 |
<0.1 |
0.5 |
– |
53.0 |
|
1975 |
17.3 |
– |
– |
3.5 |
1.0 |
21.5 |
62.0 |
0.1 |
0.4 |
– |
105.8 |
|
1976 |
15.5 |
– |
<0.1 |
3.5 |
0.3 |
23.1 |
74.4 |
0.4 |
2.3 |
<0.1 |
119.5 |
|
1977 |
17.8 |
– |
– |
4.8 |
0.2 |
27.5 |
95.8 |
0.5 |
3.9 |
<0.1 |
150.5 |
|
1978 |
17.7 |
– |
<0.1 |
3.6 |
0.1 |
51.6 |
118.2 |
0.4 |
4.4 |
– |
196.0 |
|
1980 |
11.6 |
– |
<0.1 |
0.8 |
– |
42.3 |
122.0 |
1.0 |
6.4 |
0.3 |
184.4 |
|
1981 |
6.0 |
– |
<0.1 |
0.1 |
– |
43.5 |
93.7 |
1.1 |
3.0 |
<0.1 |
147.4 |
Примечание: 1930–1968 гг. по: Яблонская с соавт. (1973), 1969–1980 гг. по: Андреева (1989), 1933 г. – только для Малого моря, 1957 г. – только для южной половины, 1973 г. – только для Большого моря, x – биомасса отдельных групп не определялась.
Первой причиной снижения биомассы аральского зообентоса, так же как и зоопланктона, явились вселенные рыбы. Салака, атерина и 6 видов бычков стали встречаться в море в 1957–1959 гг., а максимальной численности они достигли к 1960–1962 гг. (Карпевич, 1975).
Именно появление огромного количества новых потребителей бентоса привело к тому, что уже в 1961 г. (с этого же года началось резкое сокращение стока рек) наблюдалось значительное снижение биомассы донных организмов на глубинах до 10 м. В последующие годы заметно уменьшилась биомасса солоноватоводных «каспийских» моллюсков и личинок Chironomidae и в более глубоких районах, а с 1964 г. началось неуклонное снижение общей биомассы зообентоса в целом по морю (Яблонская с соавт., 1973).
В 1967–1968 гг. была отмечена самая низкая биомасса зообентоса за весь период наблюдений за зообентосом Арала. При общем снижении биомассы зообентоса, биомасса эвригалинных Cerastoderma и галофильных Caspiohydrobia не только не снизилась, а наоборот увеличилась, и с 1967 г. динамика биомассы солоноватоводных и пресноводных видов перестала определять динамику общей биомассы зообентоса Аральского моря, руководящая роль перешла к эвригалинным видам.
Увеличение общей биомассы зообентоса в 1968–1974 гг. связано с успешной акклиматизацией эвригалинных видов морского происхождения Nereis diversicolor и Syndosmya segmentum, которые на освоенных участках моря сразу становились преобладающими видами и быстро расселялись по акватории. Уже в 1970 г. биомасса зообентоса была на уровне, предшествующем ее снижению; определять ее стали акклиматизанты (в основном S. segmentum). Суммарная биомасса аборигенных видов в эти годы изменилась мало, но процесс выпадения пресноводных и солоноватоводных видов почти завершился.
Дальнейший рост биомассы зообентоса (1975–1981 гг.) вызван увеличением биомассы всех эвригалинных видов, обитавших к тому времени в Арале. В конце 1970-х годов была зарегистрирована средняя биомасса 196 г/м2, что в 8.5 раз превышает среднюю многолетнюю до зарегулирования стока рек. Несомненно, что увеличению биомассы зообентоса способствовало одновременное резкое сокращение численности рыб-бентофагов, отмечаемое Р.М. Лим и Е.Л. Марковой (1981). Тем более, что резкое возрастание численности и биомассы мирных видов в ответ на исключение хищников наблюдалось неоднократно (Holland et al., 1980; Person, 1981; и др.).
В Малом море в 1982, 1985 и 1989–1994 гг. общая численность зообентоса изменялась от 1458 до 13490 экз./м2, биомасса – 197.3–321.1 г/м2 (табл. 3.7). Обращает на себя внимание устойчиво высокая биомасса зообентоса в Малом море в последние годы. Учитывая неодновременность взятия проб, разница в отдельные годы вряд ли выходит за пределы флюктуационных изменений. Наиболее продуктивной частью Малого моря был залив Бутакова, имевший соленость близкую к 40 ‰. При проведении расширенной съемки залива в июне 1990 г. там была зарегистрирована средняя биомасса 646.0 г/м2, при колебаниях от 255.5 до 917.2 г/м2 (Андреев, Андреева, 1991а). Еще более высокая более высокая биомасса была в заливе Бутакова в августе 1992 г. На станции 18 стандартной сетки она составила 1037 г/м2, в том числе: Cerastoderma ? isthmicum – 652, Syndosmya segmentum – 336, Nereis diversicolor – 40 и Caspiohydrobia – 9 г/м2.
Осенью 1989–1991 гг. и в августе 1992 г. численность и биомасса зообентоса в Большом море колебалась от 1898 до 4748 экз./м2 и от 108.8 до 323.7 г/м2 (табл. 3.8). Следует отметить, что биомасса Caspiohydrobia в Большом море была значительно ниже, чем в Малом.
Таблица 3.7 Средняя численность и биомасса зообентоса Малого моря (по: Андреев, Андреева, 1997а)
|
Годы |
Nereis |
Chironomidae |
Dreissena |
Cerastoderma |
Syndosmya |
Theodoxus |
Caspiohydrobia |
Прочие |
Всего |
|
Численность, экз./м2 |
|||||||||
|
Июль-август 1982 |
1025 |
1 |
7 |
166 |
2074 |
205 |
3125 |
2 |
6605 |
|
Август 1985 |
160 |
– |
– |
97 |
1091 |
22 |
60 |
28 |
1458 |
|
Август 1989 |
1183 |
1 |
– |
127 |
4120 |
– |
4620 |
– |
10051 |
|
Июнь 1990 |
1033 |
– |
– |
196 |
4605 |
– |
4192 |
– |
10026 |
|
Май 1991 |
435 |
– |
– |
125 |
3423 |
– |
1489 |
– |
5472 |
|
Май 1992 |
356 |
– |
– |
138 |
2840 |
– |
1531 |
– |
4865 |
|
Август 1992 |
798 |
– |
– |
380 |
2278 |
– |
2162 |
– |
5618 |
|
Июнь 1993 |
479 |
– |
– |
198 |
6262 |
– |
6560 |
– |
13490 |
|
Июнь 1994 |
400 |
– |
– |
66 |
3979 |
– |
6978 |
– |
11423 |
|
Биомасса, г/м2 |
|||||||||
|
Июль-август 1982 |
7.9 |
<0.1 |
0.7 |
135.1 |
167.6 |
2.6 |
7.2 |
<0.1 |
321.1 |
|
Август 1985 |
3.4 |
– |
– |
89.8 |
103.4 |
0.5 |
0.2 |
0.2 |
197.5 |
|
Август 1989 |
10.5 |
<0.1 |
– |
83.2 |
169.1 |
– |
11.6 |
– |
274.4 |
|
Июнь 1990 |
7.6 |
– |
– |
134.3 |
218.9 |
– |
11.7 |
– |
372.5 |
|
Май 1991 |
5.9 |
– |
– |
124.5 |
169.9 |
– |
6.7 |
– |
307.0 |
|
Май 1992 |
3.2 |
– |
– |
75.8 |
159.9 |
– |
3.2 |
– |
242,1 |
|
Август 1992 |
9.5 |
– |
– |
95.2 |
124.4 |
– |
6.0 |
– |
235.1 |
|
Июнь 1993 |
5.3 |
– |
– |
46.7 |
177.3 |
– |
16.7 |
– |
246.0 |
|
Июнь 1994 |
3.2 |
– |
– |
15.8 |
194.4 |
– |
14.0 |
– |
227.4 |
Таблица 3.8 Средняя численность и биомасса зообентоса Большого моря (по: Андреев, Андреева, 1997а)
<![if !supportMisalignedColumns]><![endif]>
|
Годы |
Nereis |
Cerastoderma |
Syndosmya |
Caspiohydrobia |
Прочие |
Всего |
|
|
Численность, экз./м2 |
|
||||||
|
Октябрь 1989 |
218 |
73 |
936 |
637 |
34 |
1898 |
|
|
Сентябрь 1990 |
414 |
58 |
1625 |
832 |
2 |
2931 |
|
|
Сентябрь-октябрь 1991 |
599 |
75 |
2790 |
1279 |
5 |
4748 |
|
|
Август 1992 |
929 |
42 |
2347 |
896 |
6 |
4220 |
|
|
Биомасса, г/м2 |
|
||||||
|
Октябрь 1989 |
2.1 |
43.4 |
60.9 |
1.2 |
1.2 |
108.8 |
|
|
Сентябрь 1990 |
2.7 |
33.8 |
109.0 |
1.4 |
0.3 |
147.2 |
|
|
Сентябрь-октябрь 1991 |
6.6 |
79.1 |
235.2 |
2.6 |
0.2 |
323.7 |
|
|
Август 1992 |
10.8 |
21.9 |
165.8 |
2.3 |
0.2 |
201.0 |
|
|
|
|
|
|
|
|
|
|
Таким образом, к середине 1990-х годов отмечены колоссальные изменения в донной фауне Аральского моря.
Ранее было показано (Андреева, Андреев, 1987, 1990а; Андреев, 1999), что в Аральском море под влиянием осолонения и акклиматизации немногим более чем за 20 лет произошла перестройка солоноватоводных биоценозов в биоценозы, где руководящими стали эвригалинные виды морского происхождения, ранее крайне малочисленные в открытом море или же вселенные в Арал (рис. 3.17, 3.18). Начальный период осолонения и внедрения вселенцев характеризовался наличием большого числа малых по площади биоценозов и их чрезвычайной раздробленностью. По мере стабилизации условий обитания основную часть акватории занимали 2–3 биоценоза, которые, как и до начала нарушения режима моря, характеризовались небольшим числом составляющих видов зообентоса и различались не столько по видовому составу, сколько по количественному соотношению одних и тех же видов.
<![if !vml]> <![endif]>
<![endif]>
Рис. 3.17. Замена пресноводной и солоноватоводной “каспийской” фаун на морскую эвригалинную и галофильную в донных биоценозах Аральского моря (по: Андреева, Андреев, 1987). По оси абсцисс – годы; по оси ординат – площадь, %. 1 – биоценозы с доминированием пресноводных и солоноватоводных “каспийских” видов; 2 – пресноводных, солоноватоводных “каспийских”, морских эвригалинных и галофильных; 3 – морских эвригалинных и галофильных; 4 – биоценозы, в которых зообентос не обнаружен.
Столь резкие перестройки биоценозов Аральского моря в 1960–1970-х годах при, в общем-то, небольшом увеличении солености (всего на 6 ‰) были обусловлены переходом экосистемы моря через барьер критической солености (Хлебович, 1965, 1974) или хорогалинную зону (Kinne, 1971), сдвинутую в Арале до солености 12–14 ‰ (Андреев, 1981; Андреев, Андреева, 1981, 1990а), в связи с чем при повышении солености среда оказывалась не только неблагоприятной, а вообще непригодной для жизнедеятельности пресноводных и солоноватоводных каспийских видов. С другой стороны, резкое наращивание численности эвригалинных видов морского происхождения и галофильных видов материковых вод стало возможным только после повышения солености до 12–14 ‰.
<![if !vml]> <![endif]>
<![endif]>
Рис. 3.18. Внедрение акклиматизантов в донные биоценозы Аральского моря (по: Андреева, Андреев, 1987).
По оси абсцисс – годы; по оси ординат – площадь, %. 1 – биоценозы с доминированием акклиматизантов; 2 – акклиматизантов и аборигенов; 3 – аборигенов; 4 – биоценозы, в которых зообентос не обнаружен.
Итак, к началу-середине 1990-х годов в экосистеме Аральского моря насчитывалось небольшое число эвригалинных и галофильных, в основном интродуцированных, видов при доминировании в количественном отношении единичных форм. Произошла перестройка трофической структуры зоопланктона, зообентоса и ихтиофауны, характеризующаяся практически полным отсутствием хищных форм и определяемая сохранившимся набором видов (Андреев, Андреева 1998б). Но самым существенным изменением в экосистеме Аральского моря явилось изменение типа продуцирования органического вещества и, следовательно, направление потока энергии.
До середины 1970-х годов все исследователи Аральского моря (Берг, 1908; Бенинг, 1934; Никольский, 1940; Карпевич, 1960б; Яблонская, 1960; и др.) отмечали высокую прозрачность аральской воды, обилие макрофитов почти на всей его акватории. Во второй половине 1970-х годов при еще высокой прозрачности воды (в июле 1980 г. максимальная прозрачность была 16 м) нами отмечалось резкое снижение зарослей водных растений (Андреева, 1984; Андреева, Андреев, 1987, 1990а). В начале 1990-х годов наблюдалось снижение прозрачности воды. Так, если ранее была зарегистрирована максимальная прозрачность 24 м для Малого моря и 27 м для Большого (Берг, 1908; Никольский, 1940), то в августе 1992 г. в районе западной глубоководной котловины прозрачность не превышала 9–10 м, в центральной части Большого моря – 5–7 м, в Малом море – 4 м (Андреев, 1999). Макрофиты по станциям стандартной сетки не отмечались, их заросли были сконцентрированы только в прибрежье заливов. Если ранее отмечено 37 видов макрофитов (Доброхотова, 1971), то в начале 1990-х годов – только 4 (Orlova et al., 1998).
Изменение прозрачности воды связано, вероятно, с изменением баланса биогенных элементов и процесса продуцирования первичного органического вещества в этом водоеме. Следует более подробно остановиться на вопросах происхождения и распределения органического вещества в Аральском море, проанализированных А.Ф. Карпевич (1975), опиравшейся на материалы Е.А. Яблонской (1960, 1964, 1971) и Э.А. Бервальда (1964).
Развитие фитопланктона в Аральском море всегда было ослаблено из-за недостатка биогенов в водной толще и пресса зоопланктона. Основная масса органического вещества создавалась донной растительностью, на долю которой приходилось 90 % общей биомассы растений, и которая являлась основным потребителем биогенов и поставщиком органического вещества, что являлось важнейшей особенностью экосистемы Аральского моря, отмечаемой многими авторами. На долю первичной продукции, создаваемой в пелагиали, приходилось около 10 % (Карпевич, 1975).
Массовые формы зообентоса питались в основном растительными остатками (детритом), образующимися в результате отмирания макрофитов. Существенным дополнением к их рациону были микрофитобентос и микрофлора (Яблонская, 1964, 1971; Карпевич, 1975).
По происхождению и распределению органического вещества Аральское море являлось антиподом океана и Черного моря. Наиболее мощная пищевая цепь образовывалась за счет донных форм, пелагическая же пищевая цепь была крайне ослаблена (Карпевич, 1975).
Таким образом, если ранее в продуцировании первичного органического вещества ведущую роль играли макрофиты (Яблонская, 1964, 1971; Бервальд, 1964; Карпевич, 1975), покрывавшие почти все дно моря, то в конце 1980-х – начале 1990-х годов первенство перешло к фито- и бактериопланктону и частично к микрофито- и бактериобентосу. По исследованиям начала 1990?х годов (Добрынин, Королева, 1991; Сулялина, Смуров, 1993а, б; Орлова, 1993; Orlova et al., 1998) установлено существенное (примерно, на порядок) возрастание скорости процесса первичного продуцирования по сравнению с данными для летних съемок 1966 и 1968 гг. (Новожилова, 1973) при практически равных величинах деструкции в водной толще.
К началу 1980-х годов после перехода экосистемой моря через соленость 12–14 % в связи с вымиранием большинства донных беспозвоночных произошло нарушение трофической структуры аральского зообентоса (рис. 3.19), сложившаяся трофическая структура донной фауны в большей степени определялась, в отличие от других морей, имеющимся набором видов, а не характером распределения пищи. Позднее, на рубеже солености 25 % вновь происходит обеднение видового состава зообентоса, но общий характер доминирования пищевых группировок практически не изменился (Андреев, Андреева, 1998б). В водоеме преобладали собирающие детритофаги, заселившие и не свойственные ранее им биотопы. Трофическая структура донной фауны продолжала определяться имеющимся набором видов.
К началу 1990-х годов трофическая структура зоопланктона, зообентоса и ихтиофауны характеризовалась практически полным отсутствием хищных форм и определялась оставшимся набором видов. В море сформировались две почти одинаково значимые пищевые цепи: пастбищная и детритная (Андреев, Андреева, 1998б; Андреев, 1999).
<![if !vml]> <![endif]>
<![endif]>
Рис. 3.19. Соотношение трофических группировок в зообентосе Аральского моря (по: Андреева, Андреев, 1990б с дополнениями; 1936–1968 гг. — по табличным данным из: Яблонская с соавт., 1973).
По оси абсцисс – годы; по оси ординат – биомасса, %. 1 – неподвижные сестонофаги, 2 – подвижные сестонофаги, 3 – собирающие детритофаги, 4 – прочие.
Итак, в условиях квазистационарного режима экосистеме Аральского моря была присуща трансформация вещества и энергии в основном через детритную донную пищевую цепь. В последующем при осолонении моря происходило выпадение ряда трофических группировок и даже звеньев пищевых цепей. Одновременно наблюдалось изменение баланса биогенных элементов (Гидрометеорология …, 1990). К началу 1990-х годов отмечено максимальное содержание биогенных элементов в водах Аральского моря за весь период наблюдений (см. рис. 3.10–3.13), что при выпадении почти всех макрофитов способствовало в последующем резкому возрастанию скорости первичного продуцирования органического вещества в пелагиали.
Таким образом, в результате экологического кризиса, охватившего Аральское море, произошла перестройка структурно-функциональных связей, изменилось направление трансформации вещества и энергии в его экосистеме.