
Проблемы Эволюции |
О взрывном видообразовании, протекающем на наших глазах в высыхающем Аральском море. Омск: Изд-во Омского государственного педагогического университета, 2003. 382 с.
Работ по систематике Cerastoderma Аральского моря немного. Это публикации А.А. Остроумова (1907), С.А. Сидорова (1929), Я.И. Старобогатова (1974).
А.А. Остроумов (1907) указывает, по драгировочным материалам Л.С. Берга, для Аральского моря 1 вид Cardium (=Cerastoderma) edule L., подробно останавливаясь на характеристике ребер.
С.А. Сидоров (1929) выделяет среди аральских сердцевидок две основные группы: 1) типичную форму – Cardium edule L. и 2) вариетет ее – Cardium edule var. lamarcki Reeve, а также отмечает один нетипичный экземпляр, предполагая, что это новый вариетет Cardium lamarcki Reeve.
Я.И. Старобогатов (1974) в «Атласе беспозвоночных Аральского моря» – издании, завершающем исследования фауны Аральского моря при его квазистационарном режиме, приводит 2 вида Cerastoderma. Первый вид – Cerastoderma lamarcki (Reeve), представленный в Аральском море подвидом Cerastoderma lamarcki lamarcki (Reeve), обитающим также в Каспии, Азовском, Черном и Средиземном морях и вдоль атлантических берегов Западной Европы. При этом Я.И. Старобогатов (1974) не исключает возможности обособления арало-каспийской формы в самостоятельный подвид. Второй вид – Cerastoderma umbonatum (Wood), известный также из Каспия, северных и северо-восточных районов Азовского моря, из Средиземного моря и из соленых озер Африканского побережья.
До выхода в свет «Атласа…» (1974) исследователи бентоса Аральского моря объединяли эти виды под общим названием Cardium edule L. (Никитинский, 1933; Бенинг, 1934, 1935; Хусаинова, 1958; Деньгина, 1959а; Яблонская, 1960; и др.).
По последним номенклатурным изысканиям (Кафанов, 1980), для Cerastoderma lamarcki lamarcki (Reeve) предложено приоритетное название Cerastoderma rhomboides rhomboides (Lam.), для Cerastoderma umbonatum (Wood) – Cerastoderma isthmicum Issel.
Имеющееся в литературе указание (Russel, 1971) на обитание в Аральском море Cerastoderma glaucum (Bruguiere), вероятно, подразумевает обитание здесь вида отличного от Cerastoderma edule L., поскольку до выхода «Атласа…» (1974) все исследователи приводили для этого водоема именно C. edule.
Таким образом, в истекшем столетии род Cerastoderma (Cardiidae, Cardiiformes, Bivalvia) в Аральском море был представлен двумя видами.
Cerastoderma rhomboides rhomboides (Lam., 1819) (рис. 6.1 А)
Cardium edule L. var. lamarcki Reeve Сидоров, 1929; Cerastoderma lamarcki lamarcki (Reeve) Старобогатов, 1974.
По Я.И. Старобогатову (1974: 243): «Раковина овальная, овально-треугольная или овально-четырехугольная, обычно относительно тонкостенная, с мало выступающими макушками, заметно сдвинутыми к переднему концу. Спинной край мало изогнут, так что при переходе к переднему и заднему краям обычно заметны сильно закругленные углы. Поверхность покрыта широкими несколько уплощенными (особенно близ брюшного края) ребрами, в задней части раковины более раздвинутыми, чем в передней и средней. Поверхность ребер, особенно передних, покрыта поперечными чешуйками. Замочная площадка обычно довольно узкая; зубы тонкие. Задний кардинальный зуб правой створки имеет вид уплощенного в поперечном сечении клина. Нимфа довольно широкая, на заднем конце косо срезанная. Окраска бурая или буро-фиолетовая, часто с широкими концентрическими полосами, особенно в верхней трети высоты раковины. Длина до 55 мм».
<![if !vml]>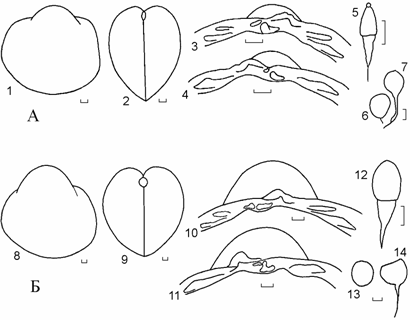 <![endif]>
<![endif]>
Рис. 6.1. Cerastoderma Аральского моря. На А – C. r. rhomboides, лагуна на острове Толмачева, 1938 г. (сборы Н.К. Алексеева); на Б – C. isthmicum, район острова Акпетки, 1936 г. (сборы Н.К. Алексеева). 1, 8 – вид сбоку; 2, 9 – вид сзади; 3, 10 – замок правой створки; 4, 11 – замок левой створки; 5, 12 – лигамент; 6,13 – отпечатки передних замыкателей; 7, 14 – отпечатки задних замыкателей. Масштаб 1 мм.
Cerastoderma isthmicum Issel, 1869 (рис. 6.1 Б.).
Cardium edule L. Сидоров, 1929, Cerastoderma umbonatum (Wood): Старобогатов, 1974.
По Я.И. Старобогатову (1974: 244): «Раковина округлая, овальная или овально-треугольная, обычно толстостенная, более вздутая, чем у предыдущего вида, с сильно выступающими макушками, сдвинутыми к переднему концу.
Спинной край равномерно изогнут так, что углы при переходе к переднему и заднему краям часто почти незаметны. Поверхность покрыта широкими, несколько расставленными округлыми в сечении ребрами, размещенными относительно равномерно. Поверхность ребер, особенно передних, покрыта частыми поперечными чешуйками. Замочная площадка очень узкая, зубы относительно тонкие. Задний кардинальный зуб правой створки имеет вид овального в поперечном сечении клина. Нимфа на заднем конце резко обрывается и подходит к замочной площадке в виде круто падающей вогнутой дуги. Окраска бурая или лиловато-бурая, иногда с широкими концентрическими полосами. Длина до 30 мм».
К началу наших исследований Аральского моря живых Cerastoderma rhomboides rhomboides уже не найдено. По Я.И. Старобогатову (1974), C. r. rhomboides обитал в прибрежных районах Арала до глубины 10–12 м, а C. isthmicum был распространен в осолоненных районах – култуках. Анализ всех имеющихся материалов позволил нам (Андреев, Андреева, 1990а, 1997а) высказать предположение, что до начала осолонения в море был распространен C. r. rhomboides – сравнительно редкая форма, а в осолоненных заливах и прилегающих к ним районам обитал C. isthmicum. По мере осолонения моря численность C. r. rhomboides снижалась, C. isthmicum проникал в открытое море. Еще в сборах по стандартной сетке станций за май 1965 г. C. r. rhomboides найден на станциях 10, 33, 42, 77, C. isthmicum – на станции 14.
В сборах 1976 г. отмечается только C. isthmicum. Наличие хорошо сохранившихся уродливых створок и раковин с лигаментом C. r. rhomboides в зоне свала глубоководной впадины в конце 1970-х годов позволило сделать заключение о вымирании C. r. rhomboides при повышении солености воды Аральского моря (Андреева, 1989).
Таким образом, к началу резких изменений биоты в середине 1970?х годов в собственно Аральском море сохранился один представитель рода Cerastoderma – C. isthmicum.
Моллюски рода Cerastoderma характеризовались исследователями донного населения Арала как малочисленная и сравнительно редко встречающаяся форма (Бенинг, 1934, 1935; Хусаинова, 1958; Яблонская, 1960; и др.). Наиболее плотные поселения имелись в мелководной зоне восточного побережья, прилегающей непосредственно к осолоненным районам – култукам и в прибрежье заливов Малого моря. В целом же моллюски были единично распространены по всей акватории Арала (рис. 6.2).
В 1964 г. в ряде районов моря зарегистрировано сокращение площадей, заселенных сердцевидками, несмотря на увеличение как средней биомассы, так и биомассы на отдельных станциях, по сравнению с 1954–1957 гг. В 1966 г. большая часть северного и западного районов и ряд южных участков моря были свободны от поселений сердцевидок, в 1967 г. продолжалось опустошение моря в западном и центральных районах. В 1968 г., как и у других видов, наблюдалось восстановление ареала и повышение средней биомассы. В многоводный 1969 г. и в последующий год вновь наблюдалось опустошение большей части акватории Большого моря, сердцевидки сохранились лишь на некоторых прибрежных станциях, и только в Малом море наблюдалось повышение биомассы этих моллюсков до 40 г/м2 и более.
Для того, чтобы разобраться в количественном распределении рода, необходимо вернуться к его видовому составу. Обитающих в Аральском море, как уже указывалось выше, Cerastoderma rhomboides rhomboides и C. isthmicum обычно определяли как Cardium edule L. и относили к эвригалинному средиземноморско-атлантическому комплексу (Зенкевич, 1963; Яблонская, 1974; и др.). По Я.И. Старобогатову (1974), Cerastoderma rhomboides rhomboides обитал в прибрежных районах Арала до глубины 10–12 м, а C. isthmicum был распространен в осолоненных районах – култуках. Еще Е.А. Яблонская (1960) высказала предположение, что церастодерма находит наиболее благоприятные условия для размножения в соленых култуках восточного и юго-восточного побережий. Налицо явное разделение видов по одному из абиотических факторов – солености. Вполне реально предположить, что такое положение было и в 1954–1957 гг.: в море был распространен C. r. rhomboides – сравнительно редкая форма, а в осолоненных заливах и прилегающих к ним районам обитал C. isthmicum. В начальный период осолонения моря снижались численность, биомасса и сокращался ареал C. r. rhomboides, в то время как C. isthmicum все еще обитал в прилегающих к осолоненным заливам районам. Где-то в 1969–1971 гг. произошло перераспределение "ролей" между видами: первый сохранился лишь в более распресненных районах, а второй, в связи с дальнейшим повышением солености, постепенно расселился по всей акватории моря.
<![if !vml]>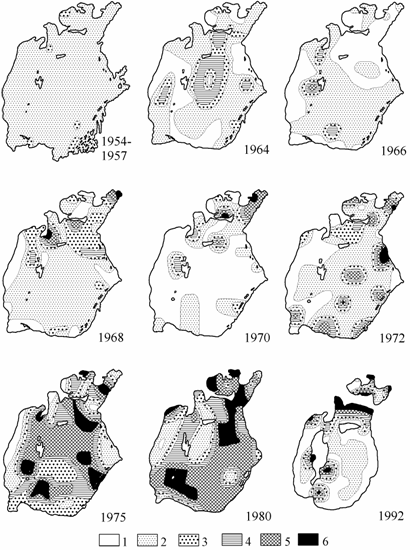 <![endif]>
<![endif]>
Рис. 6.2. Распределение биомассы Cerastoderma в Аральском море летом (по: Андреев, Андреева, 1997а; 1954–1957 гг. из: Яблонская, 1960). 1 — отсутствует, 2 — 0.1–5.0, 3 — 5.1–10.0, 4 — 10.1–20.0, 5 — 20.1–40.0, 6 — более 40 г/м2.
В пользу данного предположения свидетельствуют сохранившиеся пробы 1965 г., в которых еще в массе присутствовал C. r. rhomboides, но уже было довольно много и C. isthmicum. Далее, в период летней съемки 1976 г. в зоне свала глубоководной впадины, куда течениями сносится часть ракуши, были встречены в дночерпательных пробах в массе уродливые, хорошей сохранности, с еще заметным лигаментом створки C. r. rhomboides. Как уже отмечалось, в 1977–1980 гг. не было встречено ни одной живой особи этого вида. Скорее всего, C. r. rhomboides не приспособлен к обитанию в аральской воде повышенной солености.
На C. isthmicum – обитателе осолоненных районов – повышение солености воды моря сказалось весьма благоприятно. В 1971–1972 гг. началось активное заселение им Аральского моря, постоянное повышение его численности и биомассы (см. табл. 3.6). По мере обмеления моря и уплотнения грунта в результате накопления ракуши и хемогенных осадков проходило освоение глубоководных районов моря. Уже в 1967–1968 гг. при средней биомассе 2.1 и 3.9 г/м2 церастодерма стала давать более 20 % общей биомассы зообентоса. К концу 1970-х годов средняя биомасса церастодермы возросла до 42.3–51.6 г/м2 при максимальном значении в общей биомассе 43.6 %. Наиболее благоприятны для поселений были песчаные грунты, а также илы с большой примесью ракуши на глубинах до 20 м (Андреев, Андреева, 1997а).
В конце 1980-х – начале 1990-х годов средняя биомасса церастодермы в Малом море была более 100 г/м2, что составляло примерно 30–40 % от общей биомассы зообентоса. В Большом море биомасса сердцевидки была почти в два раза ниже и составляла около 25 % общей биомассы зообентоса (см. табл. 3.7, 3.8). На отдельных станциях Cerastoderma достигали численности 800 экз./м2 при биомассе 1482 г/м2.
Итак, на протяжении почти всего двадцатого столетия, вплоть до 1970-х годов, моллюски рода Cerastoderma в открытой части Аральского моря были редкими видами с низкой численностью и биомассой. С середины 1970-х годов отмечено резкое возрастание количественного развития Cerastoderma на фоне осолонения моря и вымирания бентосных беспозвоночных и рыб-бентофагов. В начале 1990-х годов средняя биомасса этих моллюсков превысила их среднюю биомассу до начала 1970-х годов в Большом море почти на порядок, в Малом море — на 1–2 порядка.
Расселение Cerastoderma по открытой части Аральского моря и повышение их численности сопровождались нарастанием изменчивости раковин. За период исследований Cerastoderma Аральского моря изменчивость формы раковины, ее окраски снаружи и изнутри, скульптуры, замка в целом и в строении отдельных зубов, лигамента, а также морфометрических индексов была колоссальна и подробно рассмотрена в отдельном издании (Андреева, 2000). В этой главе приведена сокращенная информация, необходимая для изложения вопросов эволюционных преобразований аральских двустворок. Особо следует отметить изменчивость скульптуры раковин и строения замка, направленную на снижение массы раковин за счет уменьшения числа и высоты ребер, утончения замочной площадки, ослабления и редукции зубов.
Общее число ребер у Cerastoderma в 1976–1994 гг. варьировало от 13 до 28 с преобладанием 19–22 ребер, нормально развитых ребер насчитывалось от 4 до 22 при преобладании 13–15. Наблюдалось смещение центра изменчивости признака в сторону уменьшения общего числа ребер на створках раковин с 21–23 в 1976–1981 гг. до 19–21 в 1994 г. и 17–19 для Cerastoderma залива Бутакова в 1993 г. Для числа нормально развитых ребер это смещение еще более выражено, так у Cerastoderma в прибрежье острова Толмачева в 1976 г. преимущественно насчитывалось 15–18 нормально развитых ребер, в Малом море в 1994 г. — 12–15, а у Cerastoderma залива Бутакова — 10–12 ребер.
Изменчивость замкового аппарата Cerastoderma за период исследований также колоссальна как по правой, так и по левой створке. Наряду с различной степенью редукции зубов наблюдались разнообразные новообразования в замке (рис. 6. 3), формирование дополнительных зубов и даже второй, а иногда и третьей нимфы (Андреева, 2000б). Спектр изменчивости замкового аппарата чрезвычайно велик, в качестве примера можно рассмотреть некоторые варианты замка левой и правой створки (рис. 6.4, 6.5). В проанализированных выборках моллюсков насчитывалось около 130 вариантов строения замка правой створки. Такие систематические признаки, как набор, величина и форма зубов, имеющие значение при дифференциации вида, рода и даже семейства у двустворок, для моллюсков семейства Cerastoderma Аральского моря во второй половине двадцатого века утратили свое значение. Изменчивы замки не отдельных особей в выборках или у моллюсков из определенных районов наблюдений, изменчивы замки всех особей на всех станциях стандартной сетки и во всех дополнительных сборах за 1989–1994 гг. В 1993–1994 гг. практически не найдено особей с типичным для какого-либо вида Cerastoderma замком. Если в редких случаях замок одной из створок более или менее соответствовал типовому описанию, то замок другой створки имел серьезные отклонения от типа.
Строение лигамента двустворчатых моллюсков является систематическим признаком (Petersen, 1958; Михайлова, 1985; и др.). У Cerastoderma Аральского моря форма, размеры и соотношение отдельных частей лигамента чрезвычайно разнообразны. Выделяются одно- и двучлениковые лигаменты со слаборазвитыми хвостообразными придатками и без продолжения впереди макушек (рис. 6.6, 161), такие же с хорошо выраженным хвостообразным придатком (рис. 6.6 256, 260), многочлениковые лигаменты с разной степенью развития хвостообразного придатка (рис. 6.6 269, 230, 255, 252 и др.). Особо следует отметить наиболее сложные по строению четковидные лигаменты, когда хвостообразный придаток развит в виде отдельных, расположенных одно за другим вздутий (рис. 6.6 277, 283, 284).
<![if !vml]>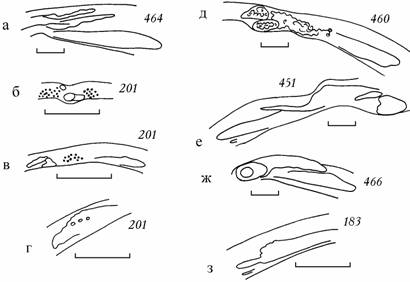 <![endif]>
<![endif]>
Рис. 6.3. Новообразования в замке Cerastoderma Аральского моря (по: Андреева, Андреев, 2000б). а – расщепление 1 заднего латерального зуба на два, б–г – жемчужные образования, д – образование зубовидных коралловых выростов на замочной площадке и кариес основания заднего латерального зуба, е –образование зубовидной пластинки из нимфы, ж – кратероподобные образования на основании 1 заднего латерального зуба, з – расщепление переднего латерального зуба. Цифры – коллекционный номер. Масштаб 1 мм.
При анализе выборок 1989–1994 гг. практически можно говорить об индивидуальных особенностях лигамента каждого отдельного моллюска. Достаточно ясно видны различия между лигаментами Cerastoderma начала исследований и последних лет. Так, моллюски 1976 г. (см. рис. 6.6 верхний ряд) имели в основном короткие, вздутые лигаменты разнообразной формы с расширенным передним или центральным участком. Число члеников варьировало без учета хвостообразного придатка от 1 до 4, очень редко достигая 6. Моллюскам из сборов 1993–1994 гг. присущи длинные, часто многочлениковые лигаменты с массивным хвостообразным придатком (см. рис. 6.6 средний и нижний ряд). Число члеников варьирует от 3 до 8, достигая в некоторых случаях 14–16. Наличие продолжения лигамента впереди макушек – морфологическая особенность отдельных особей, либо характерная особенность отдельных популяций (Андреева, 2000б). Продолжение лигамента впереди макушек может быть представлено как только слоем слияния, так и иметь типичное для двустворок (Скарлато с соавт., 1990) трехслойное строение.
<![if !vml]>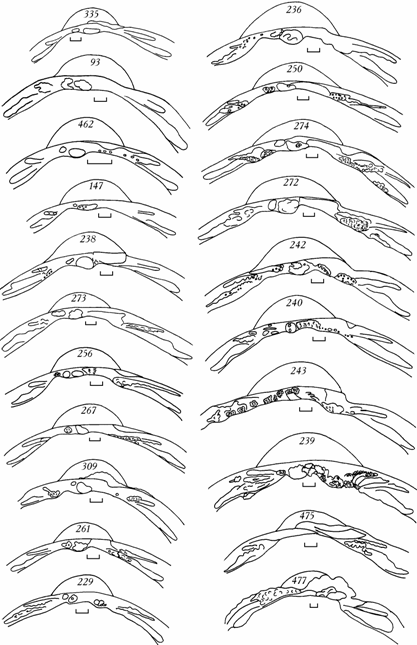 <![endif]>
<![endif]>
Рис. 6.4. Изменчивость замка правой створки Cerastoderma Аральского моря в 1976–1994 гг.
Цифры на створке – коллекционный номер. Под номером 335 – эталонный замок. Масштаб 1 мм.
<![if !vml]>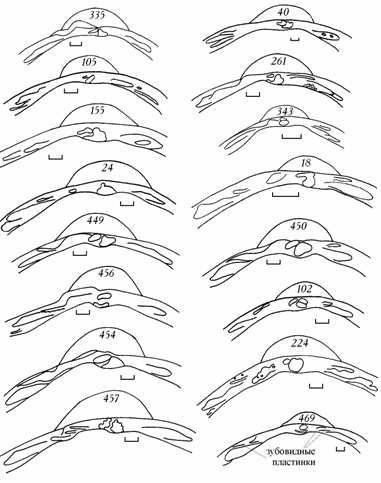 <![endif]>
<![endif]>
Рис. 6.5. Изменчивость замка левой створки Cerastoderma Аральского моря в 1976–1994 гг.
Цифры на створке – коллекционный номер. Под номером 335 – эталонный замок. Масштаб 1 мм.
<![if !vml]>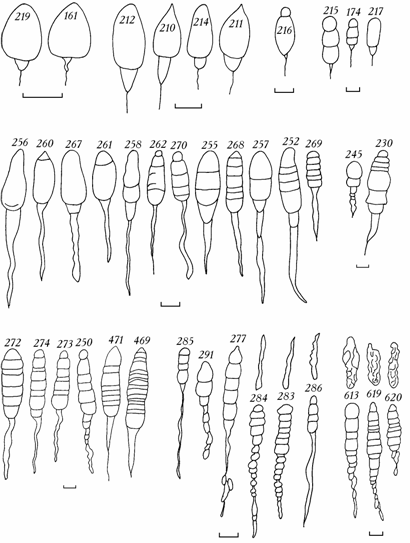 <![endif]>
<![endif]>
Рис. 6.6. Изменчивость лигамента Cerastoderma Аральского моря в 1976–1994 гг. (по: Андреева, Андреев, 2000б).
Цифры на лигаменте или над ним – коллекционный номер. Масштаб 1 мм.
Прослеживается корреляция между длиной лигамента, формой и выпуклостью раковины, строением замка. Очень выпуклым, шарообразным раковинам присущи малочлениковые короткие лигаменты без продолжения впереди макушек как в сборах 1976 г., так и 1994 г. Членистые лигаменты с мощным хвостообразным придатком и часто с продолжением лигамента впереди макушек имеются у раковин с серьезными отклонениями в строении замка и с признаками коралловости или кариеса, развитых в значительной степени. Четковидные лигаменты характерны для Cerastoderma из залива Бутакова с очень уплощенной раковиной и редуцированным первым задним латеральным зубом.
Изменчивы не только длина, высота, выпуклость и другие промеры раковин моллюсков из отдельных районов сбора, но и морфометрические индексы, как у отдельных особей из одной пробы, так и средние значения индексов из различных точек в отдельные годы. Так, высота раковины за 1976 –1994 гг. у отдельных особей составляла от 0.73 до 1.17 ее длины, выпуклость – от 0.45 до 1.03 ее длины, выпуклость – от 0.55 до 1.24 ее высоты (табл. 6.1) (ПРИМЕЧАНИЕ: ВСЕ ТАБЛИЦЫ ПОМЕЩЕНЫ В КОНЦЕ ГЛАВЫ). Средние значения индексов также изменялись существенно, выходя за пределы межвидовых различий, в частности, для азово-черноморских Cerastoderma (Скарлато, Старобогатов, 1972), и, превышая колебания отношения высоты раковины к ее длине и выпуклости раковины к ее высоте, для известных ископаемых видов Cerastoderma (Мерклин, 1974). В то же время известно, что габитуальные индексы достаточно постоянны на протяжении всей жизни моллюска, например, для Cerastoderma из Баренцева моря коэффициент вариации каждого из них не превышает 5 % (Агарова, 1979). Наибольший размах колебаний у аральских Cerastoderma имели отношение выпуклости раковины к ее длине, выпуклости раковины к ее высоте, длины макушек к длине выпуклой части лигамента и длины макушек к общей длине лигамента, при этом максимальные средние значения за разные даты наблюдений в 2–2.5 раза превышали минимальные. Наиболее уклоняющимися от эталонных по своим средним значениям были выборки из залива Сары-Чаганак со станций № 0 и № 1 1976 г., из залива Тщебас 1992 г., из залива Бутакова 1993 г. и входа в залив Бутакова 1994 г.
Итак, размах изменчивости всех исследованных признаков раковин Cerastoderma Аральского моря в начале 1990-х годов был велик, и превышал пределы изменчивости раковин современных азово-черноморских Cerastoderma (Скарлато, Старобогатов, 1972), а также ископаемых олигоценовых Cerastoderma юга СССР (Мерклин, 1974) и позднечетвертичных Cerastoderma edule lamarcki (Reeve) (Невесская, 1965). Некоторые признаки, служащие для видовой диагностики, в сложившейся ситуации утратили свое систематическое значение.
Изменчивость раковин Cerastoderma нарастала во времени (Андреева, 2000б). К началу 1990-х годов сформировались группы моллюсков, характеризующиеся определенными признаками. Ниже приводится характеристика наиболее распространенных или наиболее уклоняющихся форм Cerastoderma.
Приступая к работе над бентосом Аральского моря в 1975–1976 гг. (Андреева, 1978; Андреев, Андреева, 1981, 1987), мы имели в открытом море и крупных заливах с хорошим водообменом с морем довольно монотипическую форму Cerastoderma. К сожалению в коллекциях были собраны в основном уклоняющиеся от типа особи. Среди сохранившихся материалов найдены несколько экземпляров характеризующих эту форму для 1976–1978 гг. Раковина моллюсков округлая или округло-овальная, толстостенная, сильно вздутая, почти шарообразная (рис. 6.7) с сильно выступающими макушками, расположенными по центру раковины, либо немного смещенными к переднему краю раковины (l1/L 0.41, 0.43, 0.48). Высота раковины составляет 0.91–1.02 ее длины, выпуклость — 0.86–1.01 длины раковины и 0.92–1.02 высоты раковины. Лигамент чаще всего малосегментный, короткий (табл. 6.2). Раковины симметричные без зияния. Спинной край изогнут так, что угол при переходе к переднему краю почти не заметен, угол при переходе к заднему краю примерно равен 95–100°.
Поверхность раковины покрыта высокими и довольно широкими ребрами, округлыми или трапециевидными в поперечном сечении. Поверхность передних ребер полностью и средних ребер в самой нижней части покрыта частыми поперечными чешуйками. Общее число ребер колебалось от 18 до 22, нормально развитых от 15 до 18, наиболее выраженное и ярко окрашенное ребро 14, 15, 17 и 14–15 по счету (табл. 6.3–6.5).
<![if !vml]>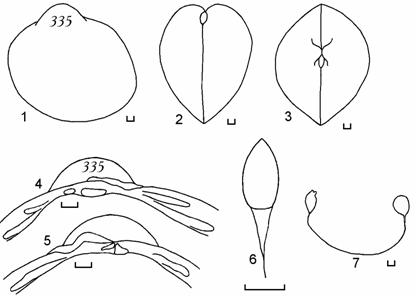 <![endif]>
<![endif]>
Рис. 6.7. Cerastoderma isthmicum (по: Андреева, Андреев, 2000в). 1 – вид сбоку, 2 – вид сзади, 3 – вид сверху, 4 – замок правой створки, 5 – замок левой створки, 6 – лигамент, 7 – отпечатки мускулов-замыкателей и мантийная линия правой створки. Цифры на створке – коллекционный номер. Масштаб 1 мм.
Из четырех эталонных экземпляров у трех обе створки имели одинаковое число ребер и одинаковое положение наиболее выраженного и ярко окрашенного ребра. У одного экземпляра наблюдалось на правой створке на два ребра больше, чем на левой. Различия в строении створок должны считаться не типичным для аральских церастодерм признаком, хотя в последующие годы при дальнейшем осолонении это явление становится нормой.
Неширокая замочная площадка имеет типичный набор довольно мощных зубов (см. рис. 6.7). Задний кардинальный зуб правой створки, габитус которого обычно используется в качестве систематического признака (Скарлато, Старобогатов 1972; Старобогатов, 1974), обнаруживает ряд вариаций формы от округло-овального и округлого правильной формы до удлиненно-овального и удлиненно-овального, изогнутого в различной степени, из-за чего для данного отрезка времени утрачивает для аральских церастодерм систематическую значимость. Кардинальные зубы левой створки свободны.
Окраска варьирует от чисто белой до желто-коричневой с белым в различных пропорциях и коричнево-бурой. У эталонных экземпляров не просматривается дефектов раковины.
Нормой реакции признаков в морфологии C. isthmicum для 1976 г. будем считать признаки эталонных экземпляров, то есть это будут моллюски с шарообразной, сильно вздутой, толстостенной раковиной с выступающими макушками, с коротким простой формы лигаментом, с большим числом ребер (18–22, нормально развитых — 15–18). Эта форма прекрасно адаптирована к почти полному закапыванию на песчаных и песчано-илистых грунтах и питанию, как указывает Н.З. Хусаинова (1958), путем фильтрации из придонных слоев волы, либо взмучиванию наилка на грунте с последующей фильтрацией.
Раковина округлой, несколько неправильной формы с немного удлиненным задним краем (рис. 6.8), толстостенная, сильно вздутая, выпуклость которой составляет 0.68–0.77 длины раковины (см. табл. 6.2), с сильно выступающими макушками, расположенными почти по центру раковины, либо немного смещенными к переднему краю (l1/L – 0.43–0.49). Раковина часто имеет асимметричные створки и иногда – зияние сзади. Угол при переходе спинного края к переднему чаще всего незаметен, к заднему – 120° и более.
<![if !vml]>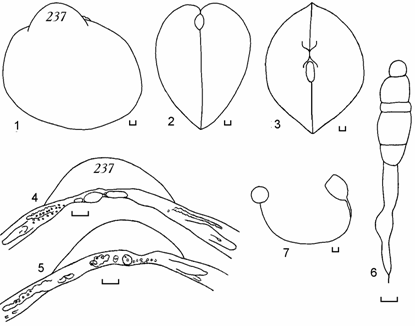 <![endif]>
<![endif]>
Рис. 6.8. Cerastoderma с округлым контуром раковины, станция 11, 1994 г. (по: Андреева, 2000а).
1 – вид сбоку, 2 – вид сзади, 3 – вид сверху, 4 – замок правой створки, 5 – замок левой створки, 6 – лигамент, 7 – отпечатки мускулов-замыкателей и мантийная линия правой створки. Цифры на створке – коллекционный номер. Масштаб 1 мм.
Поверхность раковины покрыта не очень высокими, но довольно широкими ребрами, в передней части раковины округлыми в поперечном сечении, далее трапециевидными, постепенно уплощающимися до следового отпечатка. Поверхность передних ребер и две нижние трети средних покрыты поперечными чешуйками, далее заменяемыми конхиолиновыми складочками. Общее число ребер колебалось от 19 до 21, нормально развитых – от 13 до 16, наиболее выраженное и ярко окрашенное ребро 15, 13–14, 14–15, 15–16, 16–17 по счету, а также может не отмечаться. Для большинства моллюсков отмечено различное число ребер и положение более выраженного и ярко окрашенного ребра на правой и левой створках.
Замочная площадка несколько расширена по сравнению с эталонными экземплярами Cerastoderma isthmicum 1976 г., зубы массивные, часто невысокие или уплощенные. Набор зубов типичный для C. isthmicum, но сами зубы претерпевают существенные изменения в виде расщепления латеральных, ослабления ряда зубов до рудиментарного состояния, образования на зубах коралловых и жемчужных наростов в разной степени и даже кариеса, могут присутствовать дополнительные зубовидные выросты нимфы и замочной площадки как на левой, так и на правой створке. Замок округлых, более уплощенных раковин (№ 231, 226, 225) меньше подвержен коралловости и кариесу, хотя ряд зубов может находиться в рудиментарном состоянии вплоть до полной редукции. Нимфа гладкая или с признаками коралловости и кариеса. В первом случае она широкая, короткая, с пологой выемкой на конце.
Кардинальные зубы левой створки могут быть свободными (43 %), полусросшимися (8 %) и сросшимися (49 %). Второй кардинальный правой створки округлой, часто, вследствие коралловости, неправильной формы.
Окраска раковин серо-голубая различных тонов с узкими желтыми концентрическими полосами на месте «годовых» колец, иногда с коричнево-фиолетовыми пятнами и полосами.
По сравнению с эталонными экземплярами 1976 г., эта форма обнаруживает отличия по ряду индексов (см. табл. 6.2), раковины моллюсков менее выпуклы и обладают более длинным и членистым лигаментом, в выборке встречается больше моллюсков с двумя наиболее выраженными и ярко окрашенными ребрами и меньшим числом нормально развитых ребер.
Таким образом, при сохранении внешнего облика одной жизненной формы моллюски станции 11 в 1994 г. имеют существенные отличия от эталонных 1976 г. в строении замка и лигамента, окраске створок и по другим признакам.
Раковина (рис. 6.9) толстостенная, сильно вздутая, в передней части округлая, в задней – уплощенная и довольно вытянутая с сильно выступающими макушками, расположенными почти по центру раковины, либо смещенными к переднему краю (l1/L 0.38–0.54). Примерно 35 % раковин выборки имеют асимметричные створки и зияние в задней части, иногда наблюдается зияние для ноги. Спинной край раковины изогнут таким образом, что угол при переходе к переднему краю практически незаметен, переход же к заднему краю менее плавен: примерно 35 % выборки имеют угол 120–130°, остальные 170–180°.
Поверхность раковины покрыта такими же, как у предыдущей формы, ребрами, не очень высокими, но довольно широкими, в передней части раковины округлыми в поперечном сечении, далее трапециевидными, постепенно уплощающимися до следового отпечатка. Поверхность передних и средних ребер покрыта поперечными чешуйками, далее сменяемыми конхиолиновыми складочками. От чешуек и складочек свободны обычно лишь макушки раковины. Общее число ребер колеблется от 17 до 22, число нормально развитых – от 11 до 17. Наиболее выраженное ярко окрашенное ребро 15, 17 или 13–14, 14–15 по счету, его может не быть. У большинства моллюсков отмечается одинаковое положение наиболее выраженного ярко окрашенного ребра на правой и левой створке, при различном числе ребер, причем общее число ребер правой створки, как правило, меньше на одно ребро, чем на левой.
<![if !vml]>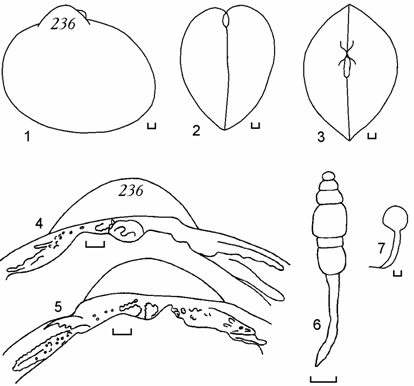 <![endif]>
<![endif]>
Рис. 6.9. Cerastoderma с немного удлиненным задним краем раковины, станция 11, 1994 г. (по: Андреева, 2000б).
1 – вид сбоку, 2 – вид сзади, 3 – вид сверху, 4 – замок правой створки, 5 – замок левой створки, 6 – лигамент, 7 – отпечаток заднего мускула-замыкателя правой створки. Цифры на створке – коллекционный номер. Масштаб 1 мм.
Замочная площадка несколько шире, чем у эталонных экземпляров C. isthmicum 1976 г., зубы массивные, часто невысокие или сглаженные. Замочная площадка имеет обычный набор зубов, часть из которых может находиться в рудиментарном состоянии, латеральные могут быть расщеплены. Большая часть зубов всех раковин в выборке имеет жемчужно-коралловые выступы и наросты, развитые в различной степени, вплоть до образования дополнительных зубовидных выростов на нимфе и зубовидных пластинок над латеральными. Примерно у половины раковин выборки замочные площадки подвержены кариесу. Нимфа может быть гладкой, шероховатой, а также с признаками коралловости и кариеса. В первом случае она широкая, удлиненная с резко обрывающимся почти под прямым углом задним краем. Кардинальные левой створки преимущественно свободны. Второй кардинальный правой створки преимущественно (70 %) округлой, часто неправильной формы, реже удлиненно-овальной. Эта группа моллюсков имеет наиболее изменчивую и сложную зубную систему, характеризующуюся образованием дополнительных зубов и зубовидных пластинок.
Раковины темного серо-голубого цвета могут иметь желто-коричневые узкие концентрические полосы, светло-серые, голубые или белые макушки, неполные концентрические полосы фиолетового цвета, распространяющиеся от задней части раковины, коричнево-фиолетовые пятна на макушках и задней части раковины.
Описываемые моллюски, по сравнению с эталонными экземплярами 1976 г. и формой с округлым контуром раковины, имеют раковину, вытянутую в длину, менее выпуклую, чем у эталонных экземпляров, с меньшим числом нормально выраженных ребер. В целом, больше зияющих, асимметричных раковин. Отмечено даже зияние для ноги.
В количественных сборах из залива Тщебас Большого моря 1992 г. обнаружено несколько групп моллюсков, имеющих раковину с очень удлиненным задним краем и различающихся по ряду морфологических признаков (строение лигамента, форма и положение макушек, скульптура и др.) и морфометрическим индексам. Одни из них близки по контуру створки к моллюскам из залива Бутакова или со станции у входа в залив Бутакова Малого моря, описываемых в следующих разделах, но отличаются строением лигамента и формой макушек, другие более сходны по всем признакам.
Анодонтоподобный и обратно мидиеподобный контуры. Раковина этих моллюсков удлиненной (анодонтоподобной или обратно мидиеподобной) формы с очень вытянутым задним краем и смещенными вперед узкими макушками, уплощенная, тонкостенная (рис. 6.10). Высота раковины составляет 0.73–0.87 ее длины, выпуклость – 0.60–0.72 длины и 0.73–0.90 высоты, у моллюсков с обратно мидиеподобным контуром – 0.80–0.82, 0.65–0.66, 0.80–0.81 соответственно (см. табл. 6.2). Лигамент мощный, членистый, имеющий морщинистое неопределенной формы продолжение впереди макушек (передний лигамент), состоящее из трех типичных слоев, а не только из слоя слияния как у моллюсков из Малого моря. Несимметричная раковина зияет сзади. Спинной край раковины изогнут так, что угол при переходе к переднему и заднему краям почти незаметен, либо в задней части раковины образуется угол примерно 170°.
Поверхность раковины покрыта широкими уплощенными ребрами в передней трети раковины округлыми в поперечном сечении, далее трапециевидными, в задней трети значительно сглаженными вплоть до следового состояния. Ребра в передней половине раковины на 2/3 покрыты поперечными чешуйками, далее переходящими в конхиолиновые складочки. Общее число ребер у моллюсков с анодонтоподобным контуром раковины колеблется от 17 до 22 при 11–16 нормально развитых, имеется два наиболее высоких, ярко окрашенных ребра (см. табл. 6.3–6.5). У моллюсков с обратно мидиеподобной формой раковины (в пробе всего 2 экз.) общее число ребер 19 и 22 при 15 и 16 нормально развитых, высокое ярко окрашенное ребро одно, 15 и 17 по счету.
Неширокая замочная площадка имеет сложную зубную систему. Правая створка имеет обычно два кардинальных зуба. Первый (передний) кардинальный зуб небольшой овальный или округлый в поперечном сечении, второй (задний) – массивный в поперечном сечении удлиненно-овальный, изогнутый в различной степени. Передние латеральные зубы, особенно второй, несколько укорочены по сравнению с задними латеральными. Первый задний латеральный зуб может быть уменьшен в различной степени, вплоть до полной редукции. Нимфа может иметь различную длину и форму, вплоть до удвоения, а также различные образования на поверхности в виде зубовидных пластинок или параллельных углублений.
<![if !vml]>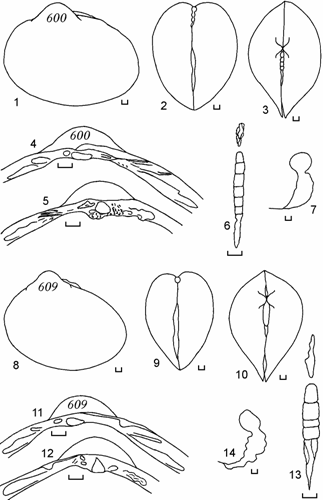 <![endif]>
<![endif]>
Рис. 6.10. Cerastoderma с очень удлиненным задним краем раковины и смещенными вперед макушками, залив Тщебас, 1992 г. (по: Андреева, 2000б). 600 – анодонтоподобная раковина, 609 – обратно мидиеподобная. 1, 8 – вид сбоку; 2, 9 – вид сзади; 3, 10 – вид сверху; 4, 11 – замок правой створки; 5, 12 – замок левой створки; 6, 13 – лигамент; 7, 14 – отпечаток заднего мускула-замыкателя правой створки. Цифры на створке – коллекционный номер. Масштаб 1 мм.
Замочная площадка левой створки еще в большей степени подвержена перестройке, она обычно подвержена коралловости и кариесу, либо же здесь возникают дополнительные зубовидные образования. Первый (передний) кардинальный зуб мощный в поперечном сечении треугольной формы, обычно окружен коралловыми образованиями. Второй (задний) кардинальный зуб разнообразной формы, часто со следами коралловости и кариеса, как и основания несколько укороченных латеральных зубов. Нимфа различной длины и формы, гладкая или с продольными параллельными невысокими ножевидными или коралловыми образованиями.
Окраска раковин коричнево-серая или светло-коричневая, изнутри – от светло-коричневой до темно-коричневой с белой перламутровой полосой по краю на заднем конце раковины.
Отпечатки передних мускулов-замыкателей типичной формы, задних – неопределенной формы. Мантийная линия в виде неполного овала, сильно расширена от отпечатков задних мускулов-замыкателей до брюшного края.
Таким образом, эти моллюски одни из наиболее уклоняющихся от эталонных 1976 г. по морфометрии раковины, строению лигамента и замкового аппарата.
Удлиненно-овальные контуры 1 и 2. Моллюски имеют удлиненно-овальную раковину с сильно вытянутым задним краем, сходны между собой по контуру створок, но различаются формой макушек и строением лигамента (рис. 6.11).
У моллюсков с удлиненно-овальным контуром 1 макушки широкие, более смещены вперед (l1/L – 0.35–0. 42, l2/L – 0.25–0.31). Лигамент длинный членистый с длинным хвостообразным придатком, имеется узкий передний лигамент, состоящий только из слоя слияния.
Моллюски с удлиненно-овальным контуром 2 имеют более узкие и высокие макушки, менее смещенные вперед (l1/L – 0.40–0.44, l2/L – 0.27–0.34). Лигамент длинный членистый с длинным членистым хвостообразным придатком и обычно широким морщинистым неопределенной формы продолжением впереди макушек типичного трехслойного строения, но с более тонкими внутренними слоями, чем собственно лигамент.
<![if !vml]>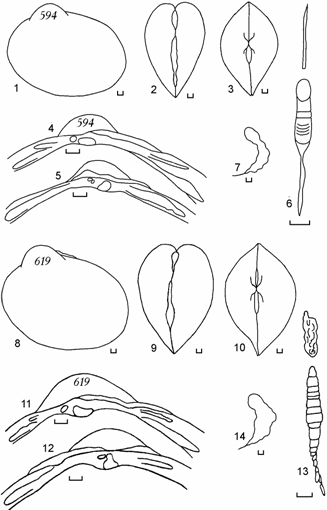 <![endif]>
<![endif]>
Рис. 6.11. Cerastoderma с очень удлиненным задним краем раковины и смещенными вперед макушками, залив Тщебас, 1992 г. (по: Андреева, 2000б). 594 – удлиненно-овальный контур 1, 619 – удлиненно-овальный контур 2. 1, 8 – вид сбоку; 2, 9 – вид сзади; 3, 10 – вид сверху; 4, 11 – замок правой створки; 5, 12 – замок левой створки; 6, 13 – лигамент; 7, 14 – отпечаток заднего мускула-замыкателя правой створки. Цифры на створке – коллекционный номер. Масштаб 1 мм.
Раковины обеих групп моллюсков тонкостенные, уплощенные (см. табл. 6.2), обычно несимметричные, с зиянием на заднем конце. Спинной край раковин у обеих групп моллюсков плавно изогнут так, что при переходе к переднему и заднему краям углы почти незаметны, либо задний угол примерно равен 170°.
Поверхность раковины покрыта плоскими ребрами, округлыми в поперечном сечении в передней трети раковины, трапециевидными во второй трети и очень плоскими, постепенно еще более уплощающимися до следового отпечатка, в задней трети. Вся раковина, за исключением выступающих частей макушек и у некоторых экземпляров примакушечной области, покрыта как бы линиями нарастания – выступающими складочками конхиолина. В передней трети раковины ребра покрыты крупными чешуйками. Общее число ребер у моллюсков с удлиненно-овальным контуром 1 колебалось от 14 до 19, нормально развитых – 9–11. Наиболее высоких и ярко окрашенных ребер может быть одно (14 или 15 по счету), либо два (11–12 и 13–14 по счету). Общее число ребер у моллюсков с удлиненно-овальным контуром 2 — 17–20 при 11–14 нормально развитых, наиболее высокого и ярко окрашенного ребра может не быть или оно одно – 13, 14, 15, 16 по счету.
У моллюсков с удлиненно-овальным контуром 1 не отмечено наружных дефектов раковин, с удлиненно-овальным контуром 2 – обычны расщепления створок по линиям нарастания.
Замочная площадка моллюсков с удлиненно-овальным контуром 1 по сравнению с эталонными экземплярами 1976 г. узкая почти без изгибов с узкими и тонкими зубами. Замок правой створки обычно имеет длинные тонкие латеральные зубы, второй передний латеральный зуб может быть укорочен. Задний (второй) кардинальный зуб правой створки массивный, удлиненно-округлый или удлиненно-овальный в поперечном сечении, иногда может быть серповидно изогнут. Нимфа очень длинная с пологой выемкой на конце.
Замок левой створки может иметь типичный набор зубов. Латеральные зубы очень длинные, кардинальные – различной формы, обычно свободны, иногда наблюдается удвоение переднего или заднего кардинального зуба. В нескольких случаях отмечена коралловость кардинальных зубов, нимфы и основания переднего латерального зуба. Нимфа длинная с пологой выемкой на конце.
Окраска раковин серо-голубая, макушки светло-коричневые. Изнутри створки коричневые, более светлого тона в передней части.
Замочная площадка моллюсков с удлиненно-овальным контуром 2 по ширине почти такая же как у эталонных экземпляров 1976 г., но более длинная и пологая с плавными изгибами.
Замок правой створки имеет обычно длинные, мощные основные латеральные зубы. Второй передний и первый задний латеральные зубы сильно ослаблены. Кардинальные зубы обычно развиты хорошо, задний (второй) кардинальный обычно серповидно изогнут. Нимфа очень длинная, как бы удвоенная, с очень пологой выемкой на конце.
Замок левой створки обычно имеет типичный набор хорошо развитых зубов. Латеральные зубы длинные, кардинальные разнообразной формы, всегда свободные. Нимфа очень длинная, как бы удвоенная, с очень пологой выемкой на конце.
Иногда, при гипертрофированном развитии лигамента и коралловости замка, может наблюдаться удвоение нимфы на задней части замочной площадки и даже формирование дополнительной нимфы в передней части замочной площадки.
Окраска раковин серо-голубая с коричневыми пятнами, макушки обычно светло-коричневые. Изнутри створки светло-коричневые, в задней половине коричневые с белой перламутровой полосой по краю створки.
Отпечатки передних мускулов-замыкателей у моллюсков обоих контуров типичной формы, задних – несколько расплывчаты и соединяются с очень расширенной мантийной линией. Мантийная линия в виде неполного овала, сильно расширена от отпечатков задних мускулов-замыкателей до брюшного края по сравнению с отрезком мантийной линии вдоль брюшного края (см. рис. 6.11 7, 14).
Таким образом, в заливе Тщебас в 1992 г. обитали моллюски, несколько сходные по внешнему облику, но имеющие существенные различия по выпуклости и скульптуре раковин, форме макушек, строению лигамента и зубной системы. Наиболее существенным различием, вероятно, следует считать формирование переднего лигамента, имеющего типичное строение, а также различную между группами, но с небольшими вариациями внутри группы скульптуру створок.
Раковина этих моллюсков удлиненной (анодонтоподобной и обратно мидиеподобной) формы с очень вытянутым задним краем и смещенными вперед широкими макушками, несколько уплощенная, с менее толстыми стенками по сравнению с эталонами 1976 г. Высота раковины у моллюсков с анодонтоподобной раковиной составляет 0.81–0.86 ее длины, выпуклость – 0.60–0.67 длины и 0.72–0.80 высоты, у обратно мидиеподобной – 0.78–0.85, 0.61–0.78 и 0.75–0.98 соответственно (см. табл. 6.2). Лигамент мощный, длинный с высоко выступающей передней частью, членистый. Раковина зияющая сзади, может иметь несимметричные створки. Спинной край раковины изогнут так, что угол при переходе к переднему краю почти незаметен. В задней части анодонтоподобных раковин спиной край может иметь характерный изгиб (рис. 6.12), образующий угол 170°, при опущенном вниз заднем крае угол равен 110°. В целом, изгиб спинного края и положение макушек таково, что моллюск, раскрыв створки, способен находиться в устойчивом вертикальном положении на грунте, опираясь на макушки и передний край.
Поверхность раковины в передней половине покрыта широкими уплощенными ребрами, округлыми в поперечном сечении, далее трапециевидными, постепенно уплощающимися вплоть до следового состояния, или же ребро может быть заметно только в примакушечной части. Примерно пятая часть поверхности раковины в задней части практически свободна от ребер или они прослеживаются только в примакушечной части. Ребра от переднего края на 1/2–1/3 длины раковины покрыты поперечными чешуйками, далее заменяемыми конхиолиновыми складочками. От них свободны только примакушечные участки. Общее число ребер колеблется от 18 до 21 при 11–14 нормально развитых ребрах у анодонтоподобных раковин и от 18 до 21 при 9–14 нормально развитых у обратно мидиеподобных. Положение наиболее выраженного и ярко окрашенного ребра различно у обеих форм, у трети особей его может не быть. Примерно у половины моллюсков отмечено различное число ребер и положение ярко окрашенного ребра на правой и левой створках.
<![if !vml]>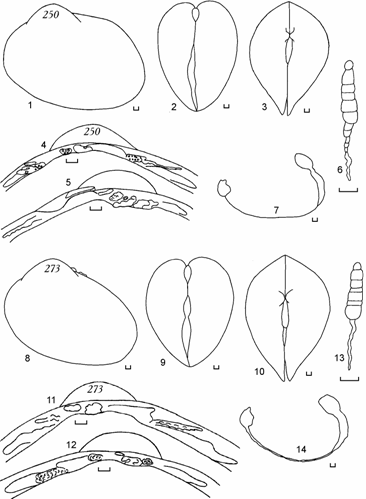 <![endif]>
<![endif]>
Рис. 6.12. Cerastoderma с очень удлиненным задним краем раковины и смещенными вперед макушками, вход в залив Бутакова, 1994 г. (по: Андреева, 2000б). 250 – анодонтоподобная раковина, 273 – обратно мидиеподобная. 1, 8 – вид сбоку; 2, 9 – вид сзади; 3, 10 – вид сверху; 4, 11 – замок правой створки; 5, 12 – замок левой створки; 6, 13 – лигамент; 7, 14 – отпечатки мускулов-замыкателей и мантийная линия правой створки. Цифры на створке – коллекционный номер. Масштаб 1 мм.
Неширокая замочная площадка имеет сложную зубную систему. Правая створка несет обычно два кардинальных зуба различного размера и формы, в разной степени подверженных коралловости и кариесу. Второй передний латеральный зуб обычно редуцирован. Его местоположение на замочной площадке и основание первого переднего латерального зуба обычно подвержены в разной степени кариесу. Первый задний латеральный зуб обычно редуцирован, и, как и основание второго заднего латерального зуба, в той или иной степени подвержен кариесу. Нимфа обычно гладкая, узкая, довольно длинная, разнообразной формы. В типичном случае рабочими остаются лишь вершины основных латеральных зубов и отчасти кардинальные зубы.
Замочная площадка левой створки еще в большей степени подвержена перестройке. Так, она в совокупности с кардинальными и передними латеральными зубами обычно подвержена коралловости и кариесу, либо же здесь возникают дополнительные зубовидные образования. Нимфа обычно гладкая, узкая, довольно длинная, разнообразной формы, может образовывать дополнительную тонкую зубовидную пластинку над задним кардинальным зубом, основание которого может быть подвержено кариесу. Кардинальные зубы левой створки могут быть как сросшимися, так и свободными. В целом, следует отметить, что эти моллюски имеют сложный зубной аппарат, дополняемый конхиолиновыми нитями, прошивающими замочные площадки обеих створок, препятствующими разъединению створок.
Окраска раковины варьирует от однотонной темно-серой до серо-голубой с желтыми узкими концентрическими полосами и коричнево-фиолетовыми пятнами на макушках и в задней части раковины, а также светло-голубой с белыми пятнами в передней части и серо-голубыми макушками и задней частью.
Мантийная линия в виде неполного овала, расширена в области сифонов, не доходит до конца задней части створки.
Таким образом, эти моллюски одни из наиболее уклоняющихся от эталонных 1976 г. как по окраске, морфометрии раковины, строению лигамента и замкового аппарата, так и по положению в грунте.
В соединящемся с морем довольно узким мелководным (глубины около 0.5 м) проливом, крупном, длиною примерно 50 км, шириною от 5 до 15 км заливе Бутакова процесс осолонения и сопутствующие ему процессы метаморфизации ионного состава воды шли опережающими темпами. Так, в июне 1990 г. в заливе зарегистрирована соленость 34.3–35.0 ‰, в прилегающих районах Малого моря 26.1 – 26.4 ‰ (Андреев, 1991). Сформировался крупный по площади изолят со своеобразным и довольно устойчивым режимом (по крайней мере в течение нескольких лет), где рассматриваемые эволюционные процессы также опережали соответствующие события в других районах моря.
К 1990 г. в заливе Бутакова сформировалась своеобразная популяция церастодермы, резко отличающаяся по габитусу и ряду морфологических признаков от обитавших ранее в море церастодерм. Раковина этих моллюсков имеет удлиненно-овальный контур 1 с сильно вытянутым задним краем (рис. 6.13.), иногда опущенным вниз, с мало выступающими макушками, смещенными к переднему краю раковины (l1/L – 0.34–0.46, l2/L – 0.25–0.31). Раковины тонкостенные, уплощенные: выпуклость раковины составляет 0.45–0.62 ее длины и 0.55–0.73 ее высоты (см. табл. 6.2), обычно симметричные с зиянием на заднем конце. Спинной край плавно изогнут так, что при переходе к заднему и переднему краям углы почти незаметны, либо задний угол примерно равен 150–170°.
<![if !vml]> <![endif]>
<![endif]>
Рис. 6.13. Cerastoderma с очень удлиненным задним краем раковины, залив Бутакова, 1993 г. (по: Андреева, 2000б). 1 – вид сбоку, 2 – вид сзади, 3 – вид сверху, 4 – замок правой створки, 5 – замок левой створки, 6 – лигамент, 7 – отпечатки мускулов-замыкателей и мантийная линия левой створки. Цифры на створке – коллекционный номер. Масштаб 1 мм.
Поверхность раковины покрыта широкими плоскими ребрами, округлыми в поперечном сечении в передней трети раковины и трапециевидными с острым углом в задней части сечения во второй трети раковины и очень плоскими, постепенно еще более уплощающимися до следового отпечатка, в задней трети. Задняя часть раковины чаще всего лишена ребер. Вся раковина за исключением выступающих частей макушек покрыта как бы линиями нарастания – выступающими складочками конхиолина. Передние ребра примерно до середины раковины покрыты крупными и довольно высокими чешуйками, размеры и форма последних способствуют парению (удержанию) моллюска в жидком илу. Общее число ребер колебалось от 14 до 19, нормально развитых – 9–14. Наиболее выраженное и ярко окрашенное ребро 14, 12–13, 13–14 и 14–15 по счету, либо его может не быть.
Особи начала 1990-х годов почти не имели дефектов раковин, в 1993–1994 гг. отмечаются небольшие искривления створок обычно в примакушечной части и небольшое расщепление раковины по линиям нарастания. До 1993 г. створки раковин были симметричны и имели обычно одинаковое число ребер и положение ярко окрашенного ребра; в популяции 1993 г. среди описываемой формы моллюсков отмечается смещение его обычно на одно ребро и различное число ребер на обеих створках.
Створки раковин соединены длинным лигаментом (длина макушек составляет 0.20–0.38 длины лигамента) с длинной выпуклой частью (длина макушек от выпуклой части лигамента составляет 0.44–0.83), часто лигамент имеет четковидную форму и продолжение (передний лигамент) впереди макушек, состоящий только из слоя слияния.
Замочная площадка узкая с узкими и тонкими зубами, имеющая существенные отличия от типовой для C. isthmicum. Замок правой створки обычно имеет сильно ослабленные второй передний и первый задний латеральные зубы и первый кардинальный зуб вплоть до их рудиментарного или следового состояния, или полной редукции первого заднего латерального зуба. Основные латеральные зубы длинные, длина переднего составляет около 1/3 от длины заднего. Задний (второй) кардинальный зуб правой створки массивный, удлиненно-округлый или удлиненно-овальный в разной степени серповидно изогнут. Нимфа длинная с полого-округлой выемкой на конце.
Замок левой створки может иметь типичный набор зубов. Задний латеральный зуб длинный, часто с маленькими зубиками у вершины – выступами замочной площадки. Латеральные зубы обычно хорошо развиты, длина переднего составляет 1/3–1/2 от длины заднего. Первый (передний) кардинальный зуб массивный, хорошо выражен, близкой к треугольной формы. Второй (задний) кардинальный зуб чаще всего редуцирован до еле заметного выступа. Кардинальные зубы обычно свободны. Нимфа длинная с полого-округлой выемкой на конце. В одном случае наблюдалось расщепление обоих латеральных зубов.
Окраска раковин темная, как у мидий, коричнево-бурая под цвет черного ила, изнутри створки окрашены в темно-коричневый цвет, по краю раковины за мантийной линией – белые с перламутровым отливом.
Отпечатки мускулов-замыкателей почти типичной формы. Мантийная линия в виде неполного овала, не заходит в заднюю часть раковины; чаще всего от отпечатков задних мускулов-замыкателей до брюшного края расширена в 3–4 раза по сравнению с отрезком мантийной линии вдоль брюшного края, либо расширена в разной степени на всем своем протяжении (см. рис. 6.13 7).
Таким образом, в заливе Бутакова сформировалась популяция моллюсков, резко отличающаяся по морфометрическим индексам от эталонных C. isthmicum 1976 г. Обитает на жидком черном иле благодаря наличию темной уплощенной зияющей раковины с очень удлиненным задним краем, меньшим числом уплощенных ребер, имеющих поперечные чешуйки и конхиолиновые складочки, с облегченной замочной площадкой за счет ослабления и редукции части зубов и усиленным развитием сифонов.
При анализе колоссальной изменчивости C. isthmicum Аральского моря, рассмотренной выше, закономерно встал вопрос о масштабах изменчивости анализируемых признаков C. isthmicum из других мест обитания и масштабах изменчивости этих же признаков у других видов Cerastoderma из ближайших к Аральскому морю водоемов. Для ответа на поставленный вопрос использованы литературные сведения (см. глава 1), материалы коллекций ЗИН РАН по Черному и Каспийскому морям и собственные сборы из береговых выбросов Азовского моря.
Cerastoderma clodiense из сборов экспедиции С.А. Зернова в августе 1911 г. (проба из хранилища ЗИН РАН, коллекционный № 48) имели слабо изменчивые раковины с удлиненно-овальным контуром створок (рис. 6.14), причем контуры створок всех присутствовавших в пробе моллюсков были практически одинаковы до такой степени, что при наложении контуры одноразмерных особей совпадали полностью, а более мелких или крупных особей казались зарисованными с одного экземпляра при разных увеличениях микроскопа. Задний край раковины несколько вытянут, широкие, мало выступающие макушки немного смещены вперед. Высота раковины составляет 0.74–0.88 ее длины, выпуклость – 0.58–0.76 длины и 0.71–0.88 высоты (табл. 6.6). Маленький, удлиненный лигамент расположен только позади макушек, одинаковой формы для всех экземпляров выборки. Симметричная раковина иногда может иметь небольшое зияние сзади. Спинной край раковины почти прямой и образует тупой угол при переходе в передний край раковины, при переходе в задний край наблюдается угол чаще всего в 130о.
Поверхность раковины покрыта гладкими трапециевидными ребрами, ширина которых незначительно превышает ширину разделяющих их промежутков, сглаженными обычно в передней и задней части раковины. Передние ребра покрыты слабо выраженными довольно тонкими чешуйками или слабыми конхиолиновыми складочками (см. рис. 1.3 3.5). Общее число ребер колеблется от 19 до 24, чаще 19 при 13–18, чаще 13 нормально развитых. Более выраженное яркоокрашенное ребро в 60 % случаев отсутствует, либо является 15, 16, 18 и 16–17 по счету.
Замочная площадка довольно узкая, слабо изогнута, почти не образует характерных для других видов Cerastoderma изгибов. Зубной аппарат как левой, так и правой створки имеет типичный набор тонких слабо изменчивых зубов. Так как в пробе имелись в основном разъединенные при предыдущих исследованиях на отдельные створки раковины, то вся изменчивость зубной системы приведена для отдельных створок (рис. 6.14 8-10). На правой створке несколько варьируют размеры второго (заднего) кардинального зуба, на левой – форма и взаиморасположение кардинальных зубов. Недлинная нимфа на заднем конце резко обрывается в виде круто падающей вогнутой дуги. В целом, зубная система C. clodiense данной выборки довольно стабильна.
<![if !vml]> <![endif]>
<![endif]>
Рис. 6.14. Cerastoderma clodiense из Черного моря (по: Андреева, 2000б).
1 – вид сбоку, 2 – вид сзади, 3 – вид сверху, 4 – лигамент, 5 – отпечаток заднего мускула-замыкателя правой створки, 6 – замок правой створки, 7 – замок левой створки, 8 – изменчивость правой створки, 9–10 – изменчивость левой створки. Цифры на створке – коллекционный номер. Масштаб 1 мм.
Окраска раковины преимущественно темная, буровато-оливковая. Изнутри раковины белые с перламутровой полосой по краю створки. Мантийная линия иногда может быть несколько расширена в области сифонов. Моллюски полностью заглублены в грунт, либо возвышаются над его поверхностью не более, чем на 1/6 длины раковины, при этом на заднем конце раковины, чуть выступающем над грунтом, расположены створки раковины усоногого рака.
В целом, выборка очень слабо изменчива, раковины почти не имеют внешних дефектов.
Cerastoderma glaucum из сборов С.А. Зернова в августе 1911 г. (проба из хранилища ЗИН РАН, коллекционный № 48) имели раковины округло-овальной или округло-четырехугольной формы. Контуры створок раковин слабо изменчивы (рис. 6.15). Задний край раковины слабо вытянут в длину. Выступающие широкие макушки несколько смещены к переднему концу раковины. Высота раковины составляет 0.83–0.91 ее длины, выпуклость – 0.64–0.73 длины и 0.75–0.86 высоты раковины (см. табл. 6.6). Маленький удлиненно-овальный лигамент несколько варьирует по форме. Раковина симметричная, обычно зияющая сзади (80 %). Спинной край раковины слабо дугообразно изогнут, при переходе к переднему и заднему краям образует закругленные углы (задний угол преимущественно 150о).
Поверхность покрыта широкими несколько уплощенными трапециевидными равномерно расставленными ребрами, промежутки между которыми заметно уже самих ребер. Поверхность ребер покрыта поперечными складочками (см. рис. 1.3 3, 5). Общее число ребер колеблется от 19 до 22, чаще 21 при 14–17, чаще 16 нормально выраженных. Более выраженное яркоокрашенное ребро (ребра) либо отсутствуют (50 %), либо являются 16, 17, 18 и 15–16, 16–17 по счету.
Относительно широкая замочная площадка слабо изогнута, зубы довольно мощные. Замковый аппарат правой створки несколько изменчив, варьирует длина и форма второго переднего латерального зуба, а также форма, размеры и положение кардинальных зубов по сравнению с типичным (см. рис. 6.15 13, 15 ,16). Вариации кардинальных зубов левой створки незначительны. Нимфа широкая, резко обрывающаяся на заднем конце. В целом, зубная система C. glaucum данной выборки довольно стабильна.
<![if !vml]> <![endif]>
<![endif]>
Рис. 6.15. Cerastoderma glaucum из Черного моря (по: Андреева, 2000б). 1 – вид сбоку, 2 – вид сзади, 3 – вид сверху, 4 – лигамент, 5 – замок правой створки, 6 – замок левой створки, 7 – отпечаток заднего мускула-замыкателя правой створки, 8 – изменчивость створок, 9–12 – изменчивость лигамента, 13, 15–16 – изменчивость правой створки, 14 – изменчивость левой створки. Цифры на створке – коллекционный номер. Масштаб 1 мм.
Окраска раковины белая или буроватая в задней части. Мантийная линия иногда может быть несколько расширена в области сифонов. Раковины не имеют внешних дефектов. Моллюски либо полностью заглублены в грунт, либо задний край раковины немного выступает над грунтом.
В целом, выборка довольно стабильна по всем изученным признакам, хотя обнаруживает несколько большую изменчивость, чем выборка C. clodiense из той же пробы.
Cerastoderma isthmicum (проба из хранилища ЗИН РАН, коллекционный № 48) экспедиции С.А. Зернова в августе 1911 г. (рис. 6.16) полностью соответствует описанию в «Определителе фауны Черного и Азовского морей» (Скарлато, Старобогатов, 1972) и сходен по морфометрическим индексам с C. isthmicum из Каспийского, Азовского морей и типовыми экземплярами из Аральского моря (см. табл. 6.6, см. табл. 6.1). Общее число ребер 20, нормально развитых – 16, более выраженного яркоокрашенного ребра нет.
Cerastoderma isthmicum из береговых выбросов Азовского моря в районе станицы Голубицкая (август 1985 г.) имели слабо изменчивые раковины с округлым контуром створок (рис. 6.17), с широкими, сильно выступающими макушками, расположенными почти посередине спинного края раковины. Спинной край раковины изогнут таким образом, что углы при переходе к переднему и заднему краям почти незаметны, либо при переходе спинного края в задний образуется угол примерно 100о. Высота раковины составляла 0.85–0.96 ее длины, выпуклость 0.66–0.98 длины и 0.76–1.02 высоты.
<![if !vml]> <![endif]>
<![endif]>
Рис. 6.16. Cerastoderma isthmicum из Черного моря (по: Андреева, Андреев, 2000д). 1 – вид сбоку, 2 – замок правой створки, 3 – замок левой створки, 4 – лигамент. Цифры на створке – коллекционный номер. Масштаб 1 мм.
<![if !vml]> <![endif]>
<![endif]>
Рис. 6.17. Cerastoderma из Азовского моря (по: Андреева, 2000б). 429 – Cerastoderma rhomboides rhomboides: 1 – вид сбоку, 2 – вид сзади, 3 – замок правой створки; 430 – Cerastoderma isthmicum: 4 – вид сбоку, 5 – вид сзади, 6 – замок правой створки. Цифры на створке – коллекционный номер. Масштаб 1 мм.
В отношении размеров лигамента и симметрии раковины судить по отдельным створкам, которыми представлена выборка, очень сложно, но, принимая во внимание стандартные, близкие к описанным в «Определителе фауны Черного и Азовского морей» (Скарлато, Старобогатов, 1972) признаки, скорее всего следует считать, что и здесь вряд ли имеются отклонения от типа.
Поверхность раковины покрыта широкими, несколько расставленными, округлыми в поперечном сечении ребрами, расположенными относительно равномерно, часто в задней части раковины сглаженными вплоть до следового состояния. Общее число ребер колеблется от 19 до 22 при 15–19 нормально развитых. Более выраженное яркоокрашенное ребро (ребра) обычно двойное 16–17 по счету, либо не наблюдается, редко 16, 17, 18–19.
Замочная площадка довольно широкая с типичным набором довольно массивных зубов. Зубной аппарат правой створки во всех случаях типичен, второй (задний) кардинальный зуб имеет форму округлого в поперечном сечении клина. Зубной аппарат левой створки типичен и лишь в одном случае наблюдалось образование небольшого дополнительного зуба над задним латеральным. Нимфа на заднем конце резко обрывается в виде круто падающей вогнутой дуги.
Окраска раковин разнообразна: от белой с коричневыми пятнами до полностью коричневой или серо-голубой. Окраска створок внутри такая же как снаружи. Мантийная линия не расширена. Примерно 40 % створок раковин имеют наружные дефекты в виде расщепления створок и вмятин на створках. В целом, это одна из наиболее стабильных по всем исследуемым признакам выборок, отвечающая типовым описаниям.
Cerastoderma rhomboides rhomboides из береговых выбросов Азовского моря в районе станицы Голубицкая (август 1985 г.) имели слабо изменчивые раковины с удлиненно-округлым контуром створок (см. рис. 6.17), с широкими, мало выступающими макушками, расположенными по центру раковины или немного смещенными вперед. Спинной край раковины мало изогнут, при переходе к переднему и заднему краям заметны сильно закругленные углы. Угол при переходе спинного края в задний чаще всего 120–130о. Высота раковины составляет 0.82–0.92 ее длины, выпуклость 0.61–0.79 длины и 0.71–0.96 высоты (см. табл. 6.6). Поскольку в выбросах присутствовали только отдельные створки, говорить о размерах лигамента и симметрии раковины не приходится.
Поверхность раковины покрыта широкими, несколько уплощенными, в задней части раковины более раздвинутыми, ребрами. Общее число ребер 18–22 при 16–19 нормально развитых. Более выраженное яркоокрашенное ребро преимущественно двойное 15–16 по счету, либо 17, 14, 14–15.
Неширокая замочная площадка имеет типичный набор довольно тонких зубов. Зубной аппарат обеих створок во всех случаях типичен, второй (задний) кардинальный зуб правой створки имеет вид овального в поперечном сечении клина. Нимфа довольно широкая, на заднем конце кососрезанная.
Окраска раковин коричнево-бурая либо серо-голубая. Внутри створки окрашены также, как и снаружи. Мантийная линия не расширена. В целом, это одна из стабильных по исследуемым признакам выборок, отвечающая типовым описаниям (Старобогатов, 1974).
Cerastoderma isthmicum из Северного Каспия в пробах за 1892 и 1897 гг. из хранилища ЗИН РАН (коллекционный № 22 и 23) имеет шаровидно вздутую раковину с овальным или удлиненно-овальным контуром (с немного удлиненным задним краем) створок, очень редко с удлиненным задним краем. Широкие, сильно выступающие макушки расположены по центру раковины либо несколько смещены вперед (рис. 6.18, 6.19). Несмотря на изменчивость контуров створок, основные морфометрические индексы имеют сравнительно небольшой размах колебаний. Высота раковины составляет 0.83–0.96 ее длины, выпуклость – 0.68–0.85 длины и 0.79–0.93 высоты (см. табл. 6.6). Небольшой лигамент разнообразен по форме, всегда двухчлениковый с округлым или удлиненным первым члеником (рис. 6.20), расположен только позади макушек. Раковина симметричная без зияния.
<![if !vml]> <![endif]>
<![endif]>
Рис. 6.18. Cerastoderma isthmicum из Каспийского моря (по: Андреева, Андреев, 2000д).
1 – вид сбоку, 2 – вид сзади, 3 – вид сверху, 4 – замок правой створки, 5 – замок левой створки, 6 – лигамент. Цифры на створке – коллекционный номер. Масштаб 1 мм.
Спинной край изогнут так, что углы при переходе к переднему и заднему краям часто почти незаметны, лишь в 44 % случаев имеется угол 110-160о при переходе спинного края в задний.
Поверхность раковины покрыта широкими несколько расставленными округлыми в поперечном сечении ребрами, расположенными относительно равномерно, часто сглаженными вплоть до следового состояния в задней части раковины. Ребра покрыты поперечными невысокими сглаженными чешуйками (см. рис. 1.3 2, 3). Общее число ребер колеблется от 16 до 21 (преимущественно 17 и 19) при 12–17 нормально выраженных (преимущественно 13). Более выраженное яркоокрашенное ребро чаще всего 12–13 либо 13 по счету, может быть 14, 15, 14–15, 15–16, его может и не быть.
<![if !vml]> <![endif]>
<![endif]>
Рис. 6.19. Изменчивость створок Cerastoderma isthmicum из Каспийского моря (по: Андреева, Андреев, 2000д).
Цифры на створке – коллекционный номер. Масштаб 1 мм.
<![if !vml]> <![endif]>
<![endif]>
Рис. 6.20. Изменчивость лигамента Cerastoderma isthmicum из Каспийского моря (по: Андреева, Андреев, 2000д).
Цифры на лигаменте – коллекционный номер. Масштаб 1 мм.
Замочная площадка широкая, довольно изогнутая с типичным набором массивных зубов. В большинстве случаев (76 %) правая и левая створки имеют типичный зубной аппарат (рис. 6.21 558, 564, 565, см. рис. 6.18), в 16 % случаев имеются незначительные отклонения от типа в виде ослабления второго переднего латерального зуба либо второго (заднего) кардинального (рис. 6.21 561, 563). И только по одному случаю (4 %) обнаружены дополнительный латеральный на левой створке (рис. 6.21 562) и проникновение лигамента внутрь замочной площадки с редукцией латеральных зубов (рис. 6.21 570). Короткая и широкая нимфа на заднем конце резко обрывается в виде круто падающей дуги. В целом, замковый аппарат C. isthmicum Каспийского моря относительно стабилен.
Окраска раковины преимущественно белая с коричневым пятном на заднем краю створки, только 8 % моллюсков в выборке имели темно-коричневые раковины. Окраска на внутренней стороне створок такая же, как и на наружной. Мантийная линия не расширена. У 20 % моллюсков наблюдались внешние дефекты створок в виде выщерблинок, искривлений створок, вмятин. Моллюски полностью заглублены в грунт, если задний конец раковины (8 % выборки) несколько выступает над грунтом, то заклеен нитями биссуса дрейссен.
В целом, выборка мало изменчива по всем анализируемым признакам.
Итак, моллюски современных видов рода Cerastoderma (C. glaucum, C. clodiense, C. isthmicum, C. rhomboides) из одного местообитания имеют слабо варьирующие морфометрические индексы и морфологические признаки.
C. isthmicum, обитавшие в Каспийском, Азовском, Черном и Аральском (сборы Л.С. Берга и типовые экземпляры 1976 г.) морях (см. табл. 6.1, 6.6), сходны по морфологическим признакам и имели близкие морфометрические индексы, хотя популяции из разных водоемов довольно различны по генотипам, о чем свидетельствуют разные для всех выборок, но относительно стабильные в пределах одной выборки, положение более выраженного яркоокрашенного ребра (ребер) на раковинах моллюсков, и, в некоторой степени, окраска раковин.
Стабильные условия обитания как в одном водоеме, так и в разных морях, способствовали отбору особей с определенным комплексом признаков, характеризующих данный вид в целом, причем разные популяции имели очень близкие, почти одинаковые морфологические признаки.
<![if !vml]> <![endif]>
<![endif]>
Рис. 6.21. Изменчивость замка Cerastoderma isthmicum из Каспийского моря (по: Андреева, Андреев, 2000д).
Цифры на правой створке – коллекционный номер, под ней левая створка. Масштаб 1 мм.
Таблица 6.1
Морфометрические индексы раковин* Cerastoderma из Аральского моря (в числителе – минимальное и максимальное значение признака, в знаменателе – среднее значение) (по: Андреева, Андреев, 2000б с дополнениями)
<![if !supportMisalignedColumns]><![endif]>
|
Место сбора и дата |
Отношение |
n |
|||||||||
|
высоты раковины к ее длине |
выпуклости раковины к ее длине |
выпуклости раковины к ее высоте |
расстояния** от переднего конца раковины до центрального ребра к длине раковины |
длины макушек к длине раковины |
длины макушек к длине выпуклой части лигамента |
длины макушек к длине лигамента |
|
||||
|
Cerastoderma isthmicum |
|||||||||||
|
Открытое море, 1900 – 1902 гг. |
0.89–0.97 0.93±0.04 |
0.71–0.96 0.83±0.13 |
0.77–1.06 0.89±0.14 |
0.48–0.52 0.50±0.02 |
0.14–0.14 0.14±0.01 |
0.95–1.00 0.98±0.30 |
_ |
5 |
|||
|
Открытое море, 1976 г. (эталонные экземпляры) |
0.91–1.02 0.95±0.08 |
0.86–1.01 0.94±0.10 |
0.92–1.02 0.98±0.07 |
0.41–0.48 0.45±0.06 |
0.13–0.21 0.16±0.06 |
0.71–1.36 1.05±0.55 |
0.43–1.13 0.74±0.52 |
|
|||
|
Прибрежье о. Толмачева, 1976 г. |
0.85–1.00 0.92±0.01 |
0.66–0.88 0.75±0.02 |
0.69–0.95 0.82±0.02 |
0.39–0.55 0.48±0.01 |
0.07–0.16 0.12±0.01 |
0.45–1.29 0.87±0.08 |
0.38–1.10 0.68±0.07 |
31 |
|||
|
Залив Сары-Чаганак, июнь 1979 г. |
0.86–0.94 0.90±0.03 |
0.73–0.79 0.77±0.02 |
0.81–0.91 0.85±0.03 |
0.39.–0.46 0.42±0.02 |
0.05–0.13 0.09±0.03 |
0.30–0.91 0.62±0.21 |
0.16–0.67 0.41±0.22 |
12 |
|||
|
Залив Сары-Чаганак, август 1979 г. |
0.76–0.98 |
0.56–1.03 |
0.59–1.24 |
0.37–0.49 |
0.08–0.16 |
0.40–1.75 |
_ |
20 |
|||
|
Станция 63, 1981 г. |
0.81–1.01 |
0.69–0.84 |
0.71–1.03 |
0.37–0.51 |
0.09–0.12 |
0.47–0.75 |
0.30–0.45 |
10 |
|||
|
Cerastoderma sp. |
|||||||||||
|
Станция 1, 1976 г. |
0.81–0.93 0.88±0.01 |
0.57–0.71 0.62±0.01 |
0.63–0.78 0.69±0.01 |
0.43–0.54 0.49±0.01 |
0.06–0.12 0.09±0.00 |
0.40–1.13 0.64±0.06 |
0.30–0.90 0.47±0.04 |
31 |
|||
Окончание табл. 6.1
<![if !supportMisalignedColumns]><![endif]>
|
Место сбора и дата |
Отношение |
n |
||||||||
|
высоты раковины к ее длине |
выпуклости раковины к ее длине |
выпуклости раковины к ее высоте |
расстояния** от переднего конца раковины до центрального ребра к длине раковины |
длины макушек к длине раковины |
длины макушек к длине выпуклой части лигамента |
длины макушек к длине лигамента |
|
|||
|
Станция 0, 1976 г. |
0.81–0.88 |
0.63–0.77 |
0.71–0.90 |
0.36–0.49 |
0.10–0.13 |
0.75–1.25 |
0.40–0.50 |
8 |
||
|
Залив Тщебас, 1992 г. |
0.73–0.87 0.82±0.01 |
0.51–0.72 0.62±0.02 |
0.64–0.90 0.76±0.02 |
0.35–0.44 0.41±0.01 |
0.08–0.14 0.10±0.01 |
0.40–0.75 0.54±0.04 |
0.21–0.48 0.33±0.03 |
26 |
||
|
Залив Бутакова, 1993 г. |
0.76–0.91 |
0.45–0.63 |
0.55–0.73 |
0.35–0.46 |
0.05–0.13 |
0.44–0.83 |
0.20–0.40 |
19 |
||
|
Район станции 11, 1994 г. |
0.75–1.17 |
0.64–0.77 |
0.65–0.95 |
0.38–0.54 |
0.08–0.19 |
0.31–0.80 |
0.29–0.64 |
27 |
||
|
Вход в залив Бутакова, 1994 г. |
0.78–0.96 0.86±0.02 |
0.53–0.77 0.64±0.02 |
0.62–0.84 0.74±0.02 |
0.37–0.50 0.43±0.01 |
0.09–0.17 0.12±0.01 |
0.55–0.93 0.71±0.04 |
0.29–0.58 0.40±0.03 |
25 |
||
|
Cerastoderma rhomboides rhomboides |
||||||||||
|
Открытое море, 1965 г. |
0.87–0.98 |
0.58–0.66 |
0.61–0.73 |
0.43–0.48 |
0.07–0.10 |
0.22–0.75 |
0.25–0.40 |
7 |
||
|
Ракуша, станция 47, 1980 г. |
0.77–0.96 0.86±0.02 |
0.52–0.86 0.69±0.04 |
0.58–1.03 0.81±0.04 |
0.35–0.50 0.40±0.01 |
0.09–0.15 0.12±0.01 |
0.71–1.11 0.84±0.06 |
0.40–0.80 0.55±0.10 |
29 |
||
Примечание: * – здесь и далее приведены 95-процентные доверительные интервалы; n – число проанализированных моллюсков, экз.; ** – измерено по продольной оси раковины; n – число проанализированных моллюсков, экз.
Таблица 6.2
Морфометрические индексы раковин Cerastoderma Аральского моря (в числителе – минимальное и
максимальное значение признака, в знаменателе – среднее значение) (по: Андреева, Андреев, 2000г)
<![if !supportMisalignedColumns]><![endif]>
|
Место сбора и дата |
Отношение |
n |
|||||||||||
|
высоты |
выпуклости раковины к ее |
расстояния* от переднего конца раковины до |
длины макушек к |
||||||||||
|
длине |
высоте |
центрального ребра к длине раковины |
середины макушек к длине раковины |
длине |
длине выпуклой части лигамента |
длине |
|||||||
Округлый контур |
|||||||||||||
|
Открытое море, 1976 г. (эталонные экземпляры) |
0.95±0.08 |
0.94±0.10 |
0.98±0.07 |
0.45±0.06 |
0.39±0.08 |
0.16±0.06 |
1.05±0.55 |
0.74±0.52 |
|
||||
|
Прибрежье о.Толмачева, 1976 г. |
0.91–0.97 0.93±0.02 |
0.71–0.88 0.78±0.04 |
0.77–0.95 0.84±0.04 |
0.39–0.49 0.46±0.02 |
0.33–0.43 0.38±0.02 |
0.10–0.14 0.13±0.01 |
0.63–1.20 0.88±0.14 |
0.47–1.10 0.77±0.15 |
10 |
||||
|
Станция 63, 1981 г. |
0.92–1.01 0.97±0.05 |
0.71–0.83 0.74±0.07 |
0.71–0.82 0.77±0.06 |
0.41-0.47 0.44±0.03 |
0.31-0.50 0.38±0.09 |
0.10–0.12 0.11±0.01 |
0.60–0.73 0.65±0.07 |
0.35–0.40 0.37±0.03 |
5 |
||||
|
Район станции 11, 1994 г. |
0.90–1.17 0.98±0.09 |
0.68–0.77 0.72±0.02 |
0.65–0.81 0.74±0.06 |
0.43–0.49 0.46±0.02 |
0.25–0.41 0.30±0.05 |
0.10–0.16 0.12±0.02 |
0.31–0.80 0.50±0.19 |
0.29–0.64 0.41±0.13 |
7 |
||||
Контур с немного удлиненным задним краем |
|||||||||||||
|
Прибрежье о.Толмачева, 1976 г. |
0.87–0.98 0.92±0.04 |
0.69–0.78 0.72±0.03 |
0.71–0.83 0.79±0.03 |
0.42–0.49 0.46±0.02 |
0.29–0.41 0.34±0.04 |
0.07–0.14 0.11±0.02 |
0.55–1.15 0.86±0.19 |
0.38–0.83 0.62±012 |
8 |
||||
|
Станция 63, 1981 г. |
0,93–0.95 0.94±0.07 |
0.69–0.73 0.71±0.21 |
0.74–0.77 0.75±0.17 |
0.44–0.46 0.45±0.13 |
0.31–0.33 0.32±0.13 |
0.09–0.11 0.10±0.14 |
0.47-0.67 0.57±1.26 |
0.30–0.33 0.31±0.24 |
2 |
||||
Продолжение табл. 6.2
<![if !supportMisalignedColumns]><![endif]>
|
Место сбора и дата |
Отношение |
n |
|||||||||||
|
высоты |
выпуклости раковины к ее |
расстояния* от переднего конца раковины до |
длины макушек к |
||||||||||
|
длине |
высоте |
центрального ребра к длине раковины |
середины макушек к длине раковины |
длине |
длине выпуклой части лигамента |
длине |
|||||||
|
Район станции 11, 1994 г. |
0.75–0.87 |
0.64–0.72 |
0.75-0.95 |
0.38–0.54 |
0.25–0.34 |
0.09–0.14 |
0.46–0.51 |
0.32–0.54 |
9 |
||||
|
Контур с очень удлиненным задним краем и смещенными макушками, в том числе: анодонтоподобный |
|||||||||||||
|
Залив Тщебас, 1992 г. |
0.73–0.87 0.82±0.04 |
0.60–0.72 0.66±0.03 |
0.73–0.90 0.81±0.05 |
0.40–0.44 0.42±0.01 |
0.28–0.35 0.31±0.02 |
0.09–0.14 0.12±0.01 |
0.46–0.75 0.57±0.08 |
0.29–0.48 0.37±0.06 |
8 |
||||
|
Вход в залив Бутакова, 1994 г. |
0.81–0.86 0.83±0.02 |
0.60–0.67 0.64±0.03 |
0.72–0.80 0.77±0.03 |
0.38–0.46 0.41±0.04 |
0.23–0.37 0.30±0.05 |
0.11–0.15 0.13±0.02 |
0.60–1.14 0.81±0.20 |
0.29–0.40 0.35±0.04 |
6 |
||||
обратно мидиеподобный |
|||||||||||||
|
Залив Тщебас, 1992 г. |
0.80–0.82 0.81±0.13 |
0.65–0.66 0.65±0.07 |
0.80–0.81 0.80±0.04 |
0.37–0.43 0.40±0.37 |
0.27–0.29 0.28±0.09 |
0.13–0.13 0.13±0.03 |
0.58–0.67 0.63±0.54 |
0.34–0.44 0.39±0.61 |
2 |
||||
|
Вход в залив Бутакова, 1994 г. |
0.78–0.85 0.80±0.03 |
0.61–0.78 0.66±0.06 |
0.75–0.98 0.83±0.09 |
0.36–0.45 0.40±0.03 |
0.29–0.35 0.31±0.03 |
0.12–0.15 0.13±0.01 |
0.60–0.86 0.67±0.10 |
0.35–0.40 0.37±0.02 |
6 |
||||
удлиненно-овальный 1 |
|||||||||||||
|
Залив Тщебас, 1992 г. |
0.76–0.84 0.80±0.03 |
0.51–0.62 0.58±0.05 |
0.64–0.80 0.73±0.07 |
0.35–0.42 0.39±0.03 |
0.25–0.31 0.29±0.03 |
0.10–0.11 0.11±0.01 |
0.54–0.75 0.64±0.11 |
0.32–0.40 0.35±0.04 |
5 |
||||
Окончание табл. 6.2
<![if !supportMisalignedColumns]><![endif]>
|
Место сбора и дата |
Отношение |
n |
|||||||||||
|
высоты |
выпуклости раковины к ее |
расстояния* от переднего конца раковины до |
длины макушек к |
||||||||||
|
длине |
высоте |
центрального ребра к длине раковины |
середины макушек к длине раковины |
длине |
длине выпуклой части лигамента |
длине |
|||||||
|
Залив Бутакова, 1993 г. |
0.76–0.87 |
0.45–0.62 |
0.55–0.73 |
0.34–0.46 |
0.25–0.31 |
0.05–0.13 |
0.44–0.83 |
0.20–0.38 |
14 |
||||
удлиненно-овальный 2 |
|||||||||||||
|
Залив Тщебас, 1992 г. |
0.79–0.86 0.82±0.01 |
0.56–0.67 0.60±0.02 |
0.68–0.79 0.74±0.02 |
0.40–0.44 0.42±0.01 |
0.27–0.34 0.31±0.02 |
0.08–0.09 0.09±0.00 |
0.40–0.51 0.46±0.03 |
0.21–0.33 0.27±0.02 |
11 |
||||
Примечание: * – измерено по продольной оси раковины;
n – число проанализированных моллюсков, экз.
Таблица 6.3
Общее число ребер на левой створке раковин «эталонных» Cerastoderma Аральского моря,
% от общего числа моллюсков (по: Андреева, 2000б)
|
Место сбора и дата |
Число ребер |
Число проанализированных створок |
|||||||||||
|
14 |
15 |
16 |
17 |
18 |
19 |
20 |
21 |
22 |
23 |
24 |
28 |
||
Округлый контур |
|||||||||||||
|
Открытое море, 1976 г. (эталонные экземпляры) |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Прибрежье о. Толмачева, 1976 г. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Станция 63, 1981 г. |
– |
– |
– |
– |
– |
– |
– |
33 |
50 |
17 |
– |
– |
6 |
|
Район станции 11, 1994 г. |
– |
– |
– |
– |
– |
29 |
14 |
57 |
– |
– |
– |
– |
7 |
Контур с немного удлиненным задним краем |
|||||||||||||
|
Прибрежье о. Толмачева, 1976 г. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Станция 63, 1981 г. |
– |
– |
– |
– |
– |
– |
– |
100 |
– |
– |
– |
– |
2 |
|
Район станции 11, 1994 г. |
– |
– |
– |
11 |
11 |
22 |
11 |
34 |
11 |
– |
– |
– |
9 |
Окончание табл. 6.3
|
Место сбора и дата |
Число ребер |
Число проанализированных створок |
|||||||||||
|
14 |
15 |
16 |
17 |
18 |
19 |
20 |
21 |
22 |
23 |
24 |
28 |
||
Контур с очень удлиненным задним краем и смещенными макушками, в том числе: |
|||||||||||||
анодонтоподобный |
|||||||||||||
|
Залив Тщебас, 1992 г. |
– |
– |
– |
25 |
25 |
38 |
– |
– |
12 |
– |
– |
– |
8 |
|
Вход в залив Бутакова, 1994 г. |
|
|
|
|
|
|
|
|
|
|
|
|
|
обратно мидиеподобный |
|||||||||||||
|
Залив Тщебас, 1992 г. |
– |
– |
– |
– |
– |
50 |
– |
– |
50 |
– |
– |
– |
2 |
|
Вход в залив |
|
|
|
|
|
|
|
|
|
– |
|
|
|
удлиненно-овальный 1 |
|||||||||||||
|
Залив Тщебас, 1992 г. |
20 |
– |
20 |
40 |
– |
20 |
– |
– |
– |
– |
– |
– |
5 |
|
Залив Бутакова, 1993 г. |
7 |
– |
29 |
14 |
43 |
7 |
– |
– |
– |
– |
– |
– |
14 |
удлиненно-овальный 2 |
|||||||||||||
|
Залив Тщебас, 1992 г. |
– |
– |
– |
20 |
20 |
30 |
30 |
– |
– |
– |
– |
– |
10 |
Таблица 6.4
Число нормальных ребер на левой створке раковин «эталонных» Cerastoderma Аральского моря,
% от общего числа моллюсков (по: Андреева, 2000б)
|
Место сбора и дата |
Число ребер |
Число проанализированных створок |
||||||||||
|
9 |
10 |
11 |
12 |
13 |
14 |
15 |
16 |
17 |
18 |
22 |
||
Округлый контур |
||||||||||||
|
Открытое море, 1976 г. (эталонные экземпляры) |
|
|
|
|
|
|
|
|
|
|
|
|
|
Прибрежье |
|
|
|
|
|
|
|
|
|
|
|
|
|
Станция 63, 1981 г. |
– |
– |
– |
– |
– |
17 |
33 |
33 |
17 |
– |
– |
6 |
|
Район станции 11, 1994 г. |
– |
– |
– |
– |
29 |
43 |
14 |
14 |
– |
– |
– |
7 |
Контур с немного удлиненным задним краем |
||||||||||||
|
Прибрежье |
|
|
|
|
|
|
|
|
|
|
|
|
|
Станция 63, 1981 г. |
– |
– |
– |
– |
– |
50 |
– |
50 |
– |
– |
– |
2 |
|
Район станции 11, 1994 г. |
– |
– |
11 |
– |
22 |
34 |
11 |
11 |
11 |
– |
– |
9 |
Окончание табл. 6.4
|
Место сбора и дата |
Число ребер |
Число проанализированных створок |
||||||||||
|
9 |
10 |
11 |
12 |
13 |
14 |
15 |
16 |
17 |
18 |
22 |
||
Контур с очень удлиненным задним краем и смещенными макушками, в том числе: |
||||||||||||
анодонтоподобный |
||||||||||||
|
Залив Тщебас, 1992 г. |
– |
– |
12 |
76 |
– |
– |
– |
12 |
– |
– |
– |
8 |
|
Вход в залив Бутакова, 1994 г. |
|
|
|
|
|
|
|
|
|
|
|
|
обратно мидиеподобный |
||||||||||||
|
Залив Тщебас, 1992 г. |
– |
– |
– |
– |
– |
– |
50 |
50 |
– |
– |
– |
2 |
|
Вход в залив Бутакова, 1994 г. |
|
|
|
|
|
|
|
|
|
|
|
|
удлиненно-овальный 1 |
||||||||||||
|
Залив Тщебас, 1992 г. |
20 |
– |
80 |
– |
– |
– |
– |
– |
– |
– |
– |
5 |
|
Залив Бутакова, 1993 г. |
14 |
14 |
29 |
36 |
– |
7 |
– |
– |
– |
– |
– |
14 |
удлиненно-овальный 2 |
||||||||||||
|
Залив Тщебас, 1992 г. |
– |
– |
20 |
10 |
30 |
40 |
– |
– |
– |
– |
– |
10 |
Таблица 6.5
Положение наиболее выраженного ярко окрашенного ребра на левой створке раковин «эталонных»
Cerastoderma Аральского моря, % от общего числа моллюсков (по: Андреева, 2000б)
<![if !supportMisalignedColumns]><![endif]>
|
Место сбора и дата |
Порядковый номер ребра ( ребер) |
Число проанализированных створок |
||||||||||||||||||||||||||
|
нет ребра |
11–12 |
12–13 |
13 |
13–14 |
14 |
14–15 |
15 |
15–16 |
16 |
16–17 |
17 |
17–18 |
18 |
22 |
||||||||||||||
Округлый контур |
||||||||||||||||||||||||||||
|
Открытое море, 1976 г. (эталонные экземпляры) |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||
|
Прибрежье |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||
|
Станция 63, 1981 г. |
– |
– |
– |
– |
16 |
– |
16 |
– |
16 |
16 |
– |
– |
– |
36 |
– |
6 |
||||||||||||
|
Район станции 11, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||
Контур с немного удлиненным задним краем |
||||||||||||||||||||||||||||
|
Прибрежье |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||
|
Станция 63, 1981 г. |
– |
– |
– |
– |
– |
– |
– |
– |
50 |
– |
– |
50 |
– |
– |
– |
2 |
||||||||||||
|
Район станции 11, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||
Окончание табл. 6.5
<![if !supportMisalignedColumns]><![endif]>
|
Место сбора и дата |
Порядковый номер ребра ( ребер) |
Число проанализированных створок |
||||||||||||||||||||||||||||
|
нет ребра |
11–12 |
12–13 |
13 |
13–14 |
14 |
14–15 |
15 |
15–16 |
16 |
16–17 |
17 |
17–18 |
18 |
22 |
||||||||||||||||
Контур с очень удлиненным задним краем и смещенными макушками, в том числе: |
||||||||||||||||||||||||||||||
анодонтоподобный |
||||||||||||||||||||||||||||||
|
Залив Тщебас, 1992 г. |
– |
– |
12 |
– |
63 |
– |
25 |
– |
– |
– |
– |
– |
– |
– |
– |
8 |
||||||||||||||
|
Вход в залив |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||
обратно мидиеподобный |
||||||||||||||||||||||||||||||
|
Залив Тщебас, 1992 г. |
– |
– |
– |
– |
– |
– |
– |
50 |
– |
– |
– |
50 |
– |
– |
– |
2 |
||||||||||||||
|
Вход в залив |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||
удлиненно-овальный 1 |
||||||||||||||||||||||||||||||
|
Залив Тщебас, 1992 г. |
– |
20 |
– |
– |
40 |
20 |
– |
20 |
– |
– |
– |
– |
– |
– |
– |
5 |
||||||||||||||
|
Залив Бутакова, 1993 г. |
7 |
– |
28 |
– |
29 |
29 |
7 |
– |
– |
– |
– |
– |
– |
– |
– |
14 |
||||||||||||||
удлиненно-овальный 2 |
||||||||||||||||||||||||||||||
|
Залив Тщебас, 1992 г. |
40 |
– |
– |
10 |
– |
20 |
– |
10 |
– |
20 |
– |
– |
– |
– |
– |
10 |
||||||||||||||
Таблица 6.6
Морфометрические индексы раковин Cerastoderma Черного, Азовского и Каспийского морей
(в числителе –минимальное и максимальное значение признака, в знаменателе – среднее значение)
<![if !supportMisalignedColumns]><![endif]>
|
Вид |
Отношение |
n |
||||||||||||||||||||
|
высоты |
выпуклости раковины к ее |
расстояния* от переднего конца раковины до |
длины макушек к |
|||||||||||||||||||
|
длине |
высоте |
центрального ребра к длине раковины |
середины макушек к длине раковины |
длине раковины |
длине выпуклой части лигамента |
длине лигамента |
||||||||||||||||
|
Черное море |
||||||||||||||||||||||
C. clodiense |
0.74–0.88 |
0.58–0.76 |
0.71–0.88 |
0.34–0.44 |
0.30–0.40 |
0.11–0.14 |
0.74–1.06 |
0.80–0.84 |
10 |
|||||||||||||
|
C. glaucum |
0.83–0.91 0.86±0.02 |
0.64–0.73 0.69±0.02 |
0.75–0.86 0.80±0.02 |
0.40–0.46 0.43±0.02 |
0.36–0.40 0.38±0.01 |
0.12–0.15 0.13±0.01 |
0.75–1.42 1.06±0.16 |
0.52–1.20 0.70±0.14 |
10 |
|||||||||||||
C. isthmicum |
0.90 |
0.72 |
0.80 |
0.45 |
0.42 |
0.15 |
1.29 |
0.88 |
1 |
|||||||||||||
|
Азовское море |
||||||||||||||||||||||
C. isthmicum |
0.85–0.96 0.90±0.02 |
0.66–0.98 0.79±0.08 |
0.76–1.02 0.88±0.06 |
0.36–0.54 0.47±0.03 |
_ |
_ |
_ |
_ |
10 |
|||||||||||||
|
C. rhomboides |
0.82–0.92 0.87±0.02 |
0.61–0.79 0.72±0.03 |
0.71–0.96 0.83±0.05 |
0.43–0.54 0.49±0.03 |
0.36–0.46 0.40±0.02 |
_ |
_ |
_ |
10 |
|||||||||||||
|
Каспийское море |
||||||||||||||||||||||
C. isthmicum |
0.83–0.96 |
0.68–0.85 |
0.79–0.93 |
0.39–0.50 |
0.34–0.45 |
0.11–0.17 |
0.71–1.38 |
0.57–1.00 |
25 |
|||||||||||||
Примечание: * – измерено по продольной оси раковины; n – число проанализированных моллюсков, экз.