
Проблемы Эволюции |
О взрывном видообразовании, протекающем на наших глазах в высыхающем Аральском море. Омск: Изд-во Омского государственного педагогического университета, 2003. 382 с.
В начале 1960-х годов в Аральское море вселен один представитель рода Syndosmya – S. segmentum Recluz.
Syndosmya segmentum Recluz, 1843
Syndesmya ovata (Phil.) Кортунова, 1970; Abra ovata (Phil.) Старобогатов, 1974.
По Я.И. Старобогатову (1974: 247): «Раковина тонкая, хрупкая, просвечивающая, равностворчатая, округло-треугольная (высота составляет 0.62–0.82 длины), уплощенная (выпуклость составляет 0.38–0.50 длины и 0.50–0.70 высоты).
Передний край округленный, задний суженный. Макушки очень узкие, мало выступающие. Поверхность покрыта тонкими линиями нарастания. Замок в правой створке состоит из двух маленьких кардинальных зубов и двух латеральных, в левой створке один маленький кардинальный зуб, латеральные не развиты.
Длина раковины до 25 мм».
К середине 1970-х годов большая часть акватории Аральского моря была заселена акклиматизантом, при этом поселения Syndosmya были чрезвычайно полиморфны по форме раковины, величине апикального угла, форме и строению лигамента, форме замочной площадки и набору зубов, конфигурации мантийной линии и отпечатков мускулов-замыкателей. Достаточно четко выделялись три формы, обитание которых было приурочено, в основном, к определенным типам грунтов (Андреева, 1980, 1999), хотя были нередки совместные находки двух каких-либо форм, а иногда и всех трех в одной дночерпательной пробе.
Подробное описание форм и рисунки приведено в разделе «Эталонные материалы Syndosmya», ниже дается таблица для их определения. Для дальнейшего описания формально обозначим их как удлиненно-овальная форма, треугольная форма и овальная форма (рис. 7.1).
Таблица для определения форм Syndosmya
1(2). Апикальный угол не менее 140о. . . . . . . . . . . . . . . .удлиненно-овальная форма.
Раковина удлиненно-овальной формы с узкими, мало выступающими макушками. Апикальный угол 140–150о. Лигамент с крупным, округлым первым сегментом, наибольшим по ширине. Примакушечный угол мантийной линии тупой и расположен не выше середины отпечатка переднего мускула-замыкателя.
2(1). Апикальный угол менее 140о.
3(4). Раковина овальной формы . . . . . . . . . . . .. . овальная форма.
Раковина овальной формы с узкими, мало выступающими макушками. Апикальный угол 130–135о. Ширина члеников лигамента одинакова на протяжении выпуклой части лигамента. Примакушечный угол мантийной линии прямой и расположен не выше верхней границы отпечатка переднего мускула-замыкателя.
4(3). Раковина треугольной формы . . . . . . . . треугольная форма.
Раковина треугольной формы с узкими, выступающими макушками. Апикальный угол 110–125о. Лигамент небольшой, узкий с маленьким, часто углубленным первым члеником. Примакушечный угол мантийной линии острый и расположен выше верхней границы отпечатка переднего мускула-замыкателя.
Нахождение удлиненно-овальной формы было приурочено, в основном, к мягким грунтам: серому и темно-серому илу центральной части моря и северных заливов, а также коричнево-серому илу в зоне влияния вод Амударьи. Обитание треугольной формы отмечено на песчаных и песчано-ракушечных грунтах, часто с зарослями зостеры, прибрежных районов моря и западной гряды островов. Встречается эта форма, хотя и в меньших количествах, на серых илах с примесью ракуши. Овальная форма найдена почти на всех типах грунтов, за исключением крупнозернистых оолитовых песков и черного ила с запахом сероводорода, однако максимального развития достигает на серых и песчанистых илах (рис. 7.2).
<![if !vml]>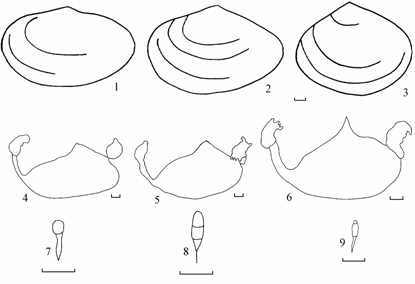 <![endif]>
<![endif]>
Рис. 7.1. Syndosmya Аральского моря (по: Андреева, Андреев, 2001а).Удлиненно-овальная форма: 1 – контур створки, 4 – контур мантийной линии и мускулов-замыкателей, 7 – лигамент; овальная форма: 2 – контур створки, 5 – контур мантийной линии и мускулов-замыкателей, 8 – лигамент; треугольная форма: 3 – контур створки, 6 – контур мантийной линии и мускулов-замыкателей, 9 – лигамент.
Во второй половине 1970-х годов не менее чем на трети станций стандартной съемки моря отмечалось совместное обитание двух каких-либо форм (при этом анализировались только крупные особи размером более 5 мм), а на трех станциях (40, 72, 93) съемки 1978 г. на сером иле были найдены одновременно в одной дночерпательной пробе все три формы Syndosmya. В последующие годы также отмечалось совместное обитание всех трех форм, в частности, на станциях 17 и в центре залива Тщебас в 1989 г. При совместном обитании всех трех форм Syndosmya максимального развития всегда достигала только одна из форм, вторая имела меньшую численность и биомассу, а третья присутствовала в небольшом количестве, как случайный элемент (табл. 7.1).
<![if !vml]>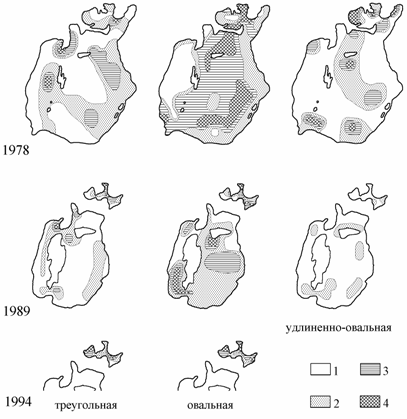 <![endif]>
<![endif]>
Рис. 7.2. Распределение биомассы форм Syndosmya Аральского моря (по: Андреева, Андреев, 2001а).
1 — отсутствует, 2 — меньше 40, 3 — 40.1–100.0, 4 — более 100 г/м2.
Таблица 7.1
Численность и биомасса форм Syndosmya при совместном обитании (по: Андреева, Андреев, 2001а)
|
Год |
Станция |
Численность, экз./м2 |
Биомасса, г/м2 |
||||
|
Удлиненно-овальная |
Треугольная |
Овальная |
Удлиненно-овальная |
Треугольная |
Овальная |
||
|
1978 |
40 |
40 |
360 |
1000 |
7.5 |
64.5 |
132.3 |
|
1978 |
72 |
190 |
20 |
530 |
45.0 |
6.7 |
114.1 |
|
1978 |
93 |
1230 |
30 |
190 |
113.9 |
12.4 |
71.3 |
|
1989 |
17 |
800 |
40 |
160 |
39.3 |
9.9 |
36.0 |
Формы Syndosmya прослеживаются с начала наших исследований над бентосом Аральского моря в 1976 г. (Андреева, 1980) вплоть до последней съемки моря в 1994 г., то есть почти на протяжении двух десятков лет при всех перестройках экосистемы моря.
Для Азовского моря отмечено, что особи S. segmentum, обитающие на ракушечнике и иле-ракушечнике, имеют более массивную и толстую раковину, чем особи, живущие в иле (Воробьев, 1949). Н.Л. Чугунов (1926) выделяет особей, живущих на ракушечных грунтах, как экологический вариетет Syndesmya ovata (=Syndosmya segmentum) f. crassa. Различия в толщине створок азовских моллюсков являются, скорее всего, проявлением биотопической изменчивости, в то время как длительное, совместное обитание в Аральском море трех форм Syndosmya, имеющих хиатус по ряду признаков, вероятно, следует рассматривать как проявление видообразовательного процесса.
Недавний акклиматизант – Syndosmya – уже в 1968 г. занял значительную часть Малого моря. По мере расселения моллюска происходило нарастание биомассы на освоенной акватории (табл. 7.2, рис. 7.3), а также в среднем по морю (см. табл. 3.2). Повышение солености совпало по времени с расселением Syndosmya и благоприятствовало ему. В 1970-х годах зафиксировано быстрое заселение моллюском дна Арала и постоянное нарастание его численности и биомассы. Количественное распределение Syndosmya в эти годы во многом сходно с распределением Cerastoderma isthmicum, с той лишь разницей, что C. isthmicum в первую очередь заселял твердые грунты, а Syndosmya, благодаря наличию различных форм (Андреева, 1980) – практически все типы грунтов (см. рис. 2.2).
Таблица 7.2
Численность и биомасса Syndosmya в июле –августе на контрольных станциях в Малом море (по: Андреева, Андреев, 2001а)
|
Год |
Численность, экз/м2 |
Биомасса, г/м2 |
||||
|
Станция 0 |
Станция 1 |
Станция 7 |
Станция 0 |
Станция 1 |
Станция 7 |
|
|
1967 |
345 |
– |
– |
14.2 |
– |
– |
|
1968 |
15 |
420 |
– |
1.8 |
51.6 |
– |
|
1969 |
5 |
115 |
80 |
0.6 |
10.6 |
16.5 |
|
1970 |
195 |
360 |
1060 |
32.4 |
92.3 |
133.1 |
|
1971 |
325 |
160 |
825 |
87.8 |
51.5 |
156.4 |
|
1972 |
45 |
275 |
875 |
17.0 |
94.5 |
142.0 |
|
1974 |
360 |
445 |
510 |
93.0 |
184.6 |
98.1 |
|
1975 |
775 |
1125 |
685 |
104.7 |
175.5 |
119.7 |
Доля Syndosmya в общей биомассе зообентоса нарастала стремительно: 1968 г. – 7.6, 1969 г. – 17.9, 1970 г. – 49.8, 1974 г. – 57.3, 1976 г. – 62.3 %. С 1974 г. значение биомассы этих моллюсков в общей биомассе зообентоса не опускалось ниже 50 %, достигнув максимума в Большом море в 1992 г. – 82.4 %. Максимальная средняя биомасса Syndosmya была зарегистрирована также в Большом море в сентябре 1991 г. – 235.2 г/м2. Высоко было значение моллюска и в общей численности, в 1970-х – начале 1980-х годов — 20.6–37.4 %, с середины 1980-х годов — 41.0–74.8 %.
Распределение биомассы форм Syndosmya (см. рис. 7.2), в основном, определялось распределением грунтов в море, что наиболее ярко прослеживается на распределении биомассы более специализированных к типам грунтов треугольной и удлиненно-овальной форм. Так, максимальная биомасса треугольной формы Syndosmya в июле 1978 г. зарегистрирована на крупнозернистых песках в западной части Большого моря и на илисто-ракушечных грунтах в Малом и вдоль восточного побережья Большого моря. Максимальная биомасса удлиненно-овальной формы Syndosmya в июле 1978 г., наоборот, была приурочена к тонкодисперсным, жидким грунтам: коричнево-серому илу в зоне влияния вод Амударьи, темно-серым и реже черным илам в центральной части Большого моря и, в основном, в заливах Малого. Менее специализированная овальная форма Syndosmya имела большую биомассу по сравнению с другими формами и больший ареал, при этом максимальная биомасса отмечена в центральных районах Малого и Большого моря с серыми, реже темно-серыми илами. Формирование овальной формой Syndosmya биомассы более 100 г/м2 на большей площади моря по сравнению с треугольной и удлиненно-овальной формами вовсе не значит, что овальная форма является доминантом в бентосе всех районов Арала. При анализе количественного распределения форм Syndosmya в 1978 г. по отдельным станциям выявлено, что на специфических биотопах все формы моллюсков этого вида развиваются в больших количествах. Так, при общей средней численности Syndosmya в июле 1978 г. 952 экз./м2 и биомассе 118.2 г/м2 треугольная форма Syndosmya на станции 63 имела численность 1230 экз./м2 и биомассу 215.4 г/м2, овальная форма Syndosmya на станции 10 – 1780 экз./м2 и 267.3 г/м2, на станции 92 – 1450 экз./м2 и 201.6 г/м2, удлиненно-овальная форма Syndosmya – на станции 1 – 3820 экз./м2 и 411 г/м2, на станции 14а – 3080 экз./м2 и 355.5 г/м2. При этом удлиненно-овальная форма, имеющая меньший по площади ареал, чем овальная форма, достигала на отдельных станциях самой большой численности и биомассы среди всех форм, превышая средние более чем в три раза.
<![if !vml]>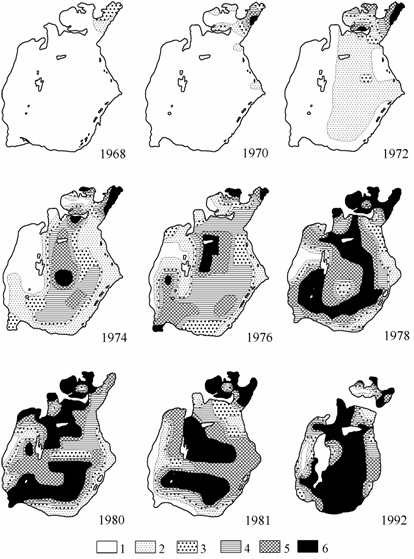 <![endif]>
<![endif]>
Рис. 7.3. Распределение биомассы Syndosmya в Аральском море летом (по: Андреев, Андреева, 1997а). 1 — отсутствует, 2 — 0.1–20.0, 3 — 20.1–40.0, 4 — 40.1–80,.0 5 — 80.1–120.0, 6 — более 120 г/м2.
Изменение характера грунтов Аральского моря, вследствие седиментации солей, к 1989 г. отрицательно сказалось на распределении удлиненно-овальной формы Syndosmya. Произошло резкое сокращение ее ареала, численности и биомассы. Ни на одной из станций не была отмечена биомасса более 40 г/м2. В 1994 г. удлиненно-овальная форма в сборах по стандартной сетке станций не обнаружена.
Для треугольной формы Syndosmya также к 1989 г. отмечено сокращение ареала, но по-прежнему ее максимальная биомасса приурочена к крупнозернистым песчаным и илисто-ракушечным грунтам, такая же ситуация сохранилась и в 1994 г.
Падение уровня моря позволило овальной форме Syndosmya освоить и часть бывших глубоководных районов, которые в 1970-х годах были непригодны для обитания моллюсков из-за специфики гидрологического режима. В то же время на некоторых станциях в северо-восточной части моря этот моллюск не был обнаружен, что, вероятно, обусловлено наличием на дне солевой корки, сцементировавшей грунты в ряде районов моря в 1989 г. В целом, распределение биомассы овальной формы Syndosmya в 1989 г. близко к таковому в 1978 г., также наиболее благоприятны для поселений центральные районы Большого моря, в Малом море максимальная биомасса отмечена в заливе Бутакова и на станциях, расположенных ближе к зоне влияния Сырдарьи. В 1994 г. свободными от поселений овальной формы были приустьевой Сырдарьинский район, а также и район глубоководной впадины, где, по данным Н.И. Андреева (1999), развивались заморные явления. Максимальная биомасса зарегистрирована в заливах и прилегающих к ним районах.
Акклиматизированный в Аральском море вид Syndosmya segmentum уже в первые годы своего обитания обнаруживает значительный полиморфизм по форме створок раковин. В наиболее ранних выборках 1970 г. отмечены моллюски, близкие по форме раковин к удлиненно-овальной форме, треугольной форме и овальной форме. Форма раковин овальной формы наиболее разнообразна, но все раковины близки к эталонным второй половины 1970-х годов. Створки раковин удлиненно-овальной формы и треугольной формы имеют переходный характер от створок овальной формы к эталонным формам второй половины 1970-х годов. В последующие годы, вероятно, отбирались моллюски, специализированные к обитанию на определенных грунтах. С 1976 г. (материалы 1971–1975 гг. на этот признак не анализировались) в Аральском море регистрируется обитание резко отличающихся по контурам створок групп моллюсков, две из которых прослеживаются до конца исследований.
Внутри каждой группы контуров обнаруживается полиморфизм (рис. 7.4–7.6), обусловленный генетической неоднородностью популяции и отчасти биотопической изменчивостью при совместном обитании разных форм. Удлиненно-овальная форма, зарегистрированная в сборах 1970, 1976–1982 и 1989–1990 гг., наибольшую изменчивость контуров створок, в том числе и уродливых, обнаруживала в сборах 1978–1980 гг. после перехода экосистемой моря барьера критической солености.
<![if !vml]>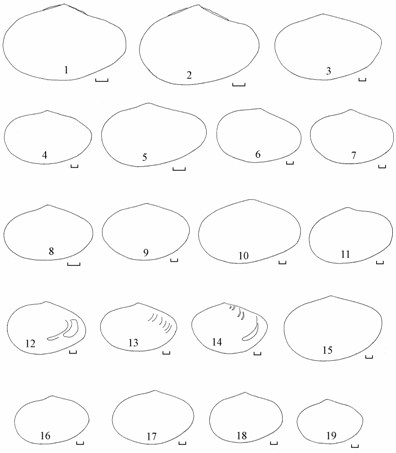 <![endif]>
<![endif]>
Рис. 7.4. Изменчивость контуров удлиненно-овальной формы Syndosmya Аральского моря (по: Андреева, Андреев, 2001а).
1–2 — 1970 г., 3–14 — 1977–1980 гг., 15–19 — 1989–1990 гг. Масштаб 1 мм.
<![if !vml]>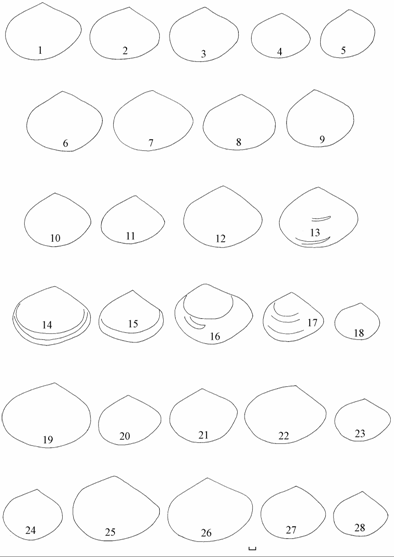 <![endif]>
<![endif]>
Рис. 7.5. Изменчивость контуров треугольной формы Syndosmya Аральского моря (по: Андреева, Андреев, 2001а). 1–5 — 1970 г., 6–19 — 1977–1980 гг., 20–28 — 1989–1994 гг. Масштаб 1 мм.
<![if !vml]>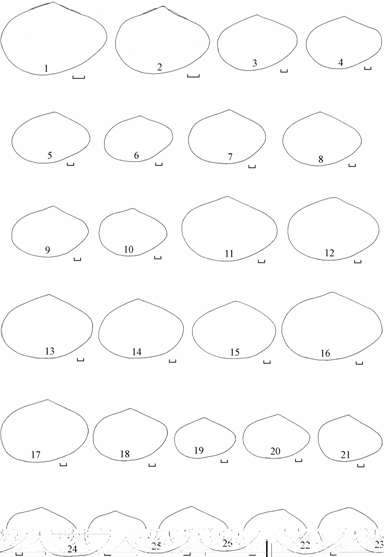 <![endif]>
<![endif]>
Рис. 7.6. Изменчивость контуров овальной формы Syndosmya Аральского моря (по: Андреева, Андреев, 2001а).
1–6 — 1970 г., 7–16 — 1977–1980 гг., 17–26 — 1989–1994 гг. Масштаб 1 мм.
У наиболее консервативной по внешнему облику треугольной формы (в сборах отмечается ежегодно) в этот период также отмечались резко уродливые створки. Большой процент уродливых створок характерен для начала 1990-х годов, когда соленость воды в ряде районов приближалась к барьерной (24–28 ‰) или же к верхней границе толерантного для Syndosmya диапазона.
Овальная форма, ежегодно отмечаемая в сборах, обнаруживала наименьшую изменчивость контуров створок вплоть до начала 1990-х годов. Только после выпадения в конце 1980-х годов солей в осадок и образования на дне плотной корки регистрируется большая вариабельность контуров и массовые микроскопические внешние дефекты раковин.
Итак, Syndosmya Аральского моря полиморфны по контурам створок раковин. Со второй половины 1970-х годов стабильно регистрируются в пробах бентоса моллюски удлиненно-овальной формы, треугольной формы и овальной формы, обитание которых приурочено к определенному типу грунта. Невооруженным глазом различия по контуру створок видны уже у особей длиною 3 мм. Моллюски внутри форм обнаруживали биотопическую изменчивость, еще более усиливавшуюся при их совместном обитании.
Строение замка Syndosmya, акклиматизированного в Аральском море, очень изменчиво во все годы наблюдений в целом и для отдельных форм. Варьируют набор, конфигурация и размеры зубов. В качестве дополнительных элементов на замочной площадке могут присутствовать различные кораллово-жемчужные образования. Четко выраженных различий в строении зубов у разных форм нет, можно лишь отметить коррелятивную зависимость общего плана строения замка от формы раковины. У удлиненно-овальной формы кардинальные зубы тонкие, маленькие, пластинчатые, латеральные зубы – тонкие, длинные. У овальной формы кардинальные и латеральные зубы хорошо развиты, не слабые. У треугольной формы кардинальные и латеральные зубы более мощные и укороченные, особенно в конце 1980-х – начале 1990-х годов, по сравнению с другими формами. Только форма замочной площадки является стабильной. У удлиненно-овальной формы замочная площадка очень пологая со слабыми изгибами перед и после ямки внутреннего лигамента, вдающейся внутрь раковины. У овальной формы замочная площадка довольно изогнута, а у треугольной формы в этой же области имеются крутые изгибы.
Ярко выраженной закономерности в изменчивости замка во временном аспекте практически нет. Так, для моллюсков удлиненно-овальной формы в 1970 г. характерно два основных набора зубов: с тонкими длинными латеральными на левой створке и без латеральных зубов на левой створке. Число латеральных зубов на правой створке – от двух до четырех, при этом форма их и размеры варьируют в широких пределах. Кардинальные зубы обеих створок, за исключением ряда экземпляров со станции 4 1970 г., маленькие, тонкие, пластинчатые. Эти два набора зубов прослеживаются у моллюсков всех выборок и в последующие годы (рис. 7.7).
В условиях критической солености появляются коралловые образования, форма и размеры кардинальных зубов значительно изменяются вплоть до полной редукции обоих кардинальных правой створки (рис. 7.7 12–13). Число, размеры и конфигурация латеральных зубов в этих условиях очень изменчивы.
При дальнейшем повышении солености наблюдается уменьшение размеров латеральных зубов на обеих створках.
У треугольной формы происходит формирование замочной площадки с характерными крутыми изгибами, которые остаются стабильным систематическим признаком для всего периода исследований. В выборках 1970 г. зарегистрированы, как и для моллюсков удлиненно-овальной формы, замочные площадки с двумя наборами зубов: с латеральными зубами на левой створке и без них (рис. 7.8). В последующем, в выборках вместо латеральных на левой створке могли присутствовать их рудименты; в выборках 1994 г. рудименты латеральных зубов на левой створке имелись лишь у единичных экземпляров, лишь на станции 15 — у половины моллюсков. Во второй половине 1970-х годов конфигурация, размеры и даже число зубов на створках значительно варьируют как у типичных, так и у уродливых экземпляров. В 1989–1990 гг. наблюдалось укорачивание основных латеральных зубов правой створки, редукция латеральных зубов на левой створке и дополнительных латеральных на правой. В выборках 1994 г. моллюски треугольной формы не имели латеральных на левой створке и дополнительных латеральных на правой, все зубы более мощные и развиты сильнее, чем у других форм (рис. 7.8 16).
<![if !vml]>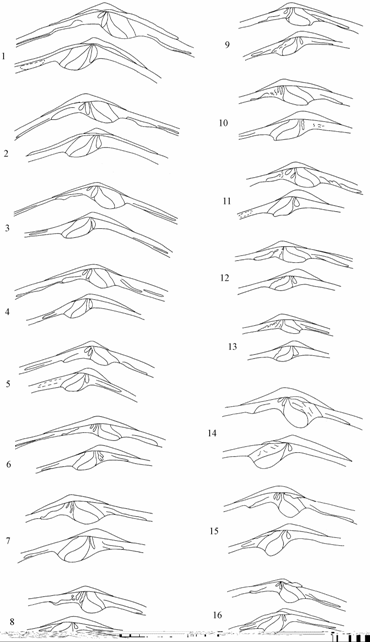 <![endif]>
<![endif]>
Рис. 7.7. Изменчивость замка удлиненно-овальной формы Syndosmya Аральского моря (по: Андреева, Андреев, 2001а).1–4 — 1970 г., 5–13 — 1977–1980 гг., 14–16 — 1989–1990 гг. Вверху правая створка, под ней левая. Масштаб 1 мм.
Для моллюсков удлиненно-овальной формы в выборке 1970 г. также отмечены замочные площадки с двумя основными наборами зубов, различающиеся наличием или отсутствием латеральных на левой створке, прослеживающиеся в течение всего периода исследований. Форма и размеры небольших, пластинчатых кардинальных зубов на обеих створках были относительно стабильны в течении всего времени наблюдений, за исключением выборки из залива Бутакова 1990 г., где моллюски обитали на жидком иле при максимальной зарегистрированной солености. Набор, размеры и конфигурация латеральных зубов обеих створок варьировали в широких пределах (рис. 7.9). В выборках 1990-х годов латеральные зубы правой створки короче и мощнее, чем в выборках предыдущих лет. Латеральные зубы левой створки в 1990-е годы у основной массы моллюсков редуцированы до небольшого бугорка в задней части замочной площадки.
В целом, замки всех Syndosmya очень изменчивы во все годы наблюдений. Проявления коралловости зарегистрированы в выборках 1970, 1980 и 1990 гг. и являются, вероятно, следствием влияния неблагоприятной солености. В 1970 г. это, скорее всего, реакция на вселение в водоем с водой, резко отличной от азовской; в 1980 г. – реакция на критическую соленость 12–14 ‰; 1990 г. – на барьерную соленость 24–28 ‰. Замки аральских Syndosmya значительно отличаются от типового описания для Syndosmya segmentum из Азовского и Черного морей (Скарлато, Старобогатов, 1972) и сходны по наличию заднего латерального зуба на левой створке и некоторым другим признакам, с замком других видов Syndosmya из Черного моря, обитающих на иных глубинах и, частично, грунтах, чем S. segmentum из того же водоема. Постоянное присутствие в выборках ряда лет у части аральских особей двух хорошо развитых латеральных зубов на левой створке и дополнительного латерального на правой выходит за пределы изменчивости замка рода Syndosmya.
Строение замка аральских Syndosmya, ввиду его колоссальной изменчивости, за исключением степени изогнутости замочной площадки, не может служить систематическим признаком.
<![if !vml]>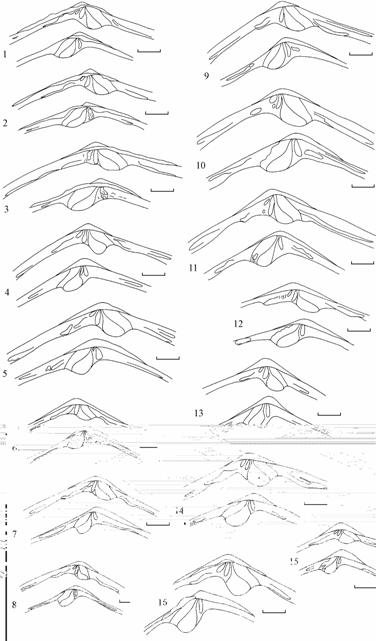 <![endif]>
<![endif]>
Рис. 7.8. Изменчивость замка треугольной формы Syndosmya Аральского моря (по: Андреева, Андреев, 2001а). 1–3 — 1970 г., 4–11 — 1977–1980 гг., 12–16 — 1989–1994 гг. Вверху правая створка, под ней левая. Масштаб 1 мм.
Форма и размеры лигамента Syndosmya, акклиматизированной в Аральском море, существенно варьируют. Наблюдается определенная тенденция изменений лигамента в зависимости от периода исследований и формы моллюска (рис. 7.10–7.12).
Наиболее сложными по строению являются лигаменты моллюсков со станции 4 сборов 1970 г., отнесенные к форме, промежуточной между овальной и треугольной. Лигаменты этих моллюсков имеют высокие, сильно развитые, членистые хвостообразные придатки и передние лигаменты. Лигаменты остальных форм в сборах 1970 г. имеют менее сложное строение, но изменчивость их очень велика. Наблюдаются лигаменты с различной членистостью основной части наружного лигамента и различным развитием хвостообразного придатка. Перед-
<![if !vml]>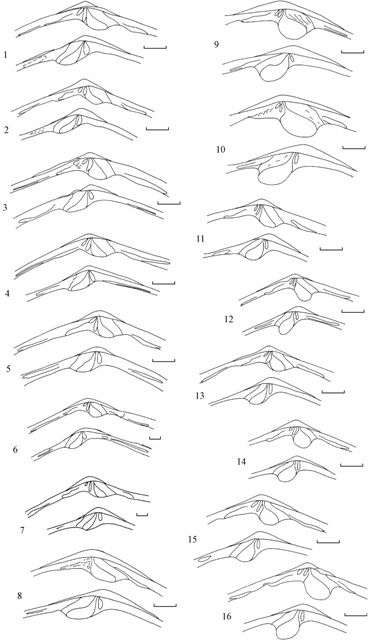 <![endif]>
<![endif]>
¬Рис. 7.9. Изменчивость замка овальной формы Syndosmya Аральского моря (по: Андреева, Андреев, 2001а).
1–4 — 1970 г., 5–10 — 1977–1980 гг., 11–16 — 1989–1994 гг. Вверху правая створка, под ней левая. Масштаб 1 мм.
<![if !vml]>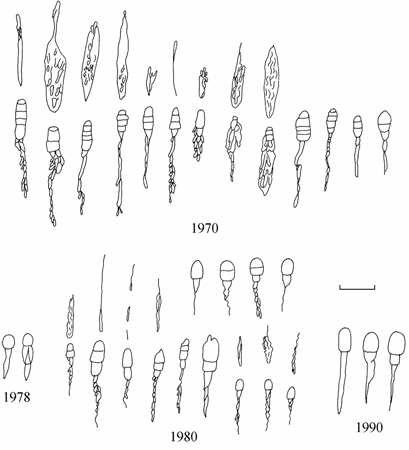 <![endif]>
<![endif]>
Рис. 7.10. Изменчивость лигамента удлиненно-овальной формы Syndosmya Аральского моря (по: Андреева, Андреев, 2001а).
<![if !vml]>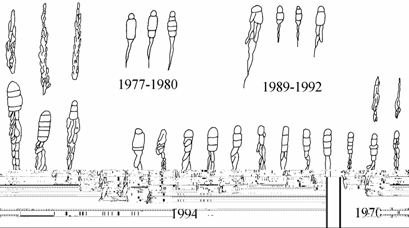 <![endif]>
<![endif]>
Рис. 7.11. Изменчивость лигамента треугольной формы Syndosmya Аральского моря (по: Андреева, Андреев, 2001а).
ний лигамент варьирует по своей форме и размерам, примерно у половины моллюсков его может не быть.
Для второй половины 1970-х годов, в основном, характерны достаточно стабильные и вполне морфологически определенные лигаменты для всех трех форм моллюсков. Для моллюсков удлиненно-овальной формы характерны одно- и двухсегментные лигаменты с массивным, чаще всего, нечленистым хвостообразным придатком. Первый членик лигамента обычно овальной формы, превышающий по ширине остальные. Для моллюсков треугольной формы характерны небольшие, узкие двух- и трехсегментные лигаменты с маленьким первым сегментом. Моллюски овальной формы имеют одно- и двухсегментные, удлиненно-овальные лигаменты с одинаковыми по ширине сегментами.
В 1980 г. вновь наблюдалась повышенная изменчивость лигамента у удлиненно-овальной и овальной форм. В 1990–1992 гг. увеличивается изменчивость лигаментов, при сохранении у большинства моллюсков внешнего облика лигамента соответствующей формы. В 1994 г. зарегистрировано резкое повышение изменчивости лигаментов у сохранившихся форм за счет увеличения членистости и разрастания хвостообразного придатка. Наибольшие вариации отмечены для треугольной формы; так, у моллюсков со станции 17 (у входа в залив Бутакова, подвергшегося наибольшему осолонению) вновь отмечено появление передних лигаментов.
Таким образом, в 1970–1994 гг. форма и размеры лигамента Syndosmya, акклиматизированных в Аральском море, имели пики повышенной изменчивости в первые годы после вселения, в 1980 г. и в последний год наблюдений.
<![if !vml]> <![endif]>
<![endif]>
Рис. 7.12. Изменчивость лигамента овальной формы Syndosmya Аральского моря (по: Андреева, Андреев, 2001а).
В период исследований внутренняя поверхность створки аральских Syndosmya ниже замочной площадки имела различное строение. Она могла быть гладкой и фарфоровидной с перламутровым отливом, с более или менее четко выделяющейся мантийной линией и отпечатками мускулов-замыкателей. Очень часто в примакушечной части створки выше примакушечного угла мантийной линии и вокруг, а иногда и на отпечатках замыкателей развивались образования в виде жемчужно-коралловых выступов, наплывов, кратеров, подобные образованиям на замочных площадках Cerastoderma (рис. 7.13). Особенно интересно образование гребня в виде ножевидного, длинного выроста, располагающегося от примакушечного угла мантийной линии до отпечатка переднего мускула-замыкателя. Интенсивность его развития может быть различной вплоть до рудимента в виде следовой линии. Утолщение створки в этой зоне может резко уменьшаться, образуя вместо гребня как бы ступеньку. Вырост или ступенька располагаются вдоль линии соприкосновения проекции на сторону нижнего края пищеварительной железы (печени) и границы прикрепления полужабр и ротовых лопастей. Жемчужно-коралловые образования служат, вероятно, для упрочнения раковины. Гребень или ступенька, а в 1994 г. и дополнительная мантийная линия, могут быть проявлениями дополнительного развития мускулатуры прикрепления полужабр и ротовых лопастей. Причем, на правой створке утолщения и гребень всегда развиты сильнее, чем на левой. Во времени и у моллюсков, принадлежащих к различным формам, эти образования развиты различным образом.
Подобные, очень слаборазвитые единичные образования у нескольких моллюсков переходных к удлиненно-овальной форме, были отмечены только в 1970 г. при заглубленном состоянии отпечатков замыкателей. В последующие годы тонкие створки удлиненно-овальной формы изнутри были гладкие со слабыми отпечатками замыкателей.
<![if !vml]>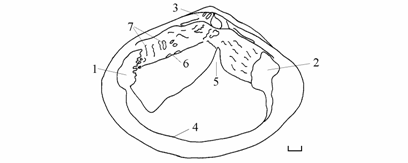 <![endif]>
<![endif]>
Рис. 7.13. Внутренняя поверхность створки Syndosmya
Аральского моря. 1 – отпечаток переднего мускула-замыкателя, 2 – отпечаток заднего мускула-замыкателя, 3 – замочная площадка, 4 – мантийная линия, 5 – примакушечный угол мантийной линии, 6 – гребень, 7 – кораллово-жемчужные образования. Масштаб 1 мм.
У моллюсков овальной формы в 1970 г. утолщения были развиты слабо либо в виде единичных выступов. Гребень был развит только у 30 % выборки в виде невысокой линии, разделяющей область проекции на сторону пищеварительной железы и область проекции полужабр и ротовых лопастей. Отпечатки замыкателей были заглублены и окружены небольшим валиком с кораллово-жемчужными образованиями. В 1977–1980 гг. у моллюсков этой формы примакушечная часть створки была утолщена в различной степени. У четверти выборки 1977-1978 гг. имелся гребень в виде острого выступа или ступеньки. В 1980 г. среди моллюсков с утолщением раковины в примакушечной части преобладают особи без четко выраженного гребня. В 1990–1992 гг. в Большом море у 60 % моллюсков, обитающих на типичных для формы серых мягких илах, подобных утолщений и развития гребня не прослеживалось. У моллюсков, обитавших вдоль восточного побережья на более плотных грунтах и при более высокой солености, в примакушечной части створок имелось утолщение, но без развития гребня. У всех моллюсков, независимо от места обитания, отмечено углубление отпечатков замыкателей и развитие в той или иной степени кораллово-жемчужных образований на границах отпечатков. В 1994 г. в Малом море только 25–30 % особей в выборках этой формы имели створки без утолщения. У большинства особей гребень не прослеживался, у единиц – в виде ступеньки. Отпечатки замыкателей у 30–50 % моллюсков в выборках были углублены и окружены коралловыми образованиями.
У моллюсков треугольной формы утолщения створок были развиты сильнее, чем у моллюсков других форм в течение всего периода наблюдений. В пробах 1970 г., когда они имели раковины с контуром, промежуточным между эталонными экземплярами треугольной и овальной форм второй половины 1970-х годов, утолщения в примакушечной части раковины были развиты в такой де степени, как и у моллюсков овальной формы из этой же выборки. В 1977–1980 гг. у всех моллюсков треугольной формы во всех выборках створки раковин были довольно толстыми с очень сильно развитым утолщением в примакушечной части, причем, не менее 90 % выборки имели гребень в аиде острого выступа или сильно развитой ступеньки. Створки раковин этой формы по толщине сравнимы со створками одноразмерных сердцевидок, обитавших на этих же станциях. У всех особей отпечатки замыкателей заглублены и окружены сильно выраженными коралловыми выростами. В 1989–1992 гг. створки раковин становятся более тонкими и хрупкими, утолщения в примакушечной части ?аввиты в виде слабых или единично расположенных наплывов. Примерно половина выборки имела гребень в виде ступеньки или следового отпечатка разной степени выраженности. Отпечатки замыкателей у всех моллюсков заглублены и окружены коралловыми наростами. В выборках 1994 г. из Малого моря сильно утолщенные створки имеют от 5 до 70 % моллюсков. Гребень, в основном, развит в виде следового отпечатка. Отпечатки замыкателей несколько заглублены и окружены коралловым выростами.
Итак, из анализа имеющихся в нашем распоряжении материалов следует, что утолщения створок в примакушечной области у моллюсков треугольной и овальной форм Аральского моря были наиболее развиты во второй половине 1970-х годов, в период, когда крупнозернистые пески занимали еще довольно обширные участки дна моря, а серые илы на многих станциях имели значительную примесь ракуши. Обнажение песчаных, песчано-илистых и илисто-ракушечных грунтов мелководных районов моря, изменение химического состава воды приводит к ослаблению толщины створок в 1989–1992 гг., в том числе и в примакушечной области. Вполне вероятно, что утолщение створок в примакушечной части обусловлено механическим составом грунтов, на которых обитали моллюски. Так, мкллюски удлиненно-овальной формы, приспособленной к обитанию на тонкодисперсных жидких черных и темно-серых илах, в наших сборах 1976–1990 гг. подобного утолщения не имели. Максимально же развиты утолщения в этой области у моллюсков треугольной формы, обитавших на крупнозернистых (оолитовых) песках станции 63 и в прибрежье острова Лазарева.
Для Syndosmya Аральского моря в сборах 1970–1994 гг. прослеживается изменение очертаний мантийной линии в зависимости от формы моллюска и периода исследований (рис. 7.14–7.16), а отпечатков мускулов-замыкателей – в зависимости от периода исследований (рис. 7.17–7.19). Так, для 1970 г., когда еще не наблюдалось достаточно четкого разграничения Syndosmya на формы, очертания мантийной линии и отпечатков замыкателей сильно варьируют. Отпечатки чаще всего одинаковы на обеих створках, хотя встречаются особи с различными размерами и контурами отпечатков на левой и правой створках. Наблюдаются увеличение извилистости границ отпечатка и изменение его формы за счет, вероятно, развития дополнительных мускульных тяжей. Это явление более характерно для переднего замыкателя моллюсков овальной формы и части особей со станции 16, более сходных с удлиненно-овальной формой. В целом, отпечатки замыкателей моллюсков станции 4 имеют более ровные границы, чем у моллюсков станции 16, за исключением особей, тяготеющих по внешнему облику к треугольной форме.
<![if !vml]> <![endif]>
<![endif]>
Рис. 7.14. Изменчивость контура мантийной линии удлиненно-овальной формы Syndosmya Аральского моря (по: Андреева, Андреев, 2001а). 1 — 1970 г., 2 — 1977 г., 3–5 — 1980 г., 6 — 1989 г. Масштаб 1 мм.
Для второй половины 1970-х годов характерны слабая изменчивость контуров мантийной линии и их специфика для каждой формы. Достаточно четко выделяется контур мантийной линии треугольной формы, имеющей острый угол в примакушечной части створки, вершина которого располагается выше верхней границы переднего замыкателя или в более редких случаях на ее уровне.
Контуры мантийной линии моллюсков удлиненно-овальной и овальной форм более сходны между собой и различаются положением угла в примакушечной части и его размерами. У удлиненно-овальной формы угол тупой и расположен он не выше середины отпечатка переднего замыкателя, в то время как у овальной формы угол прямой и расположен обычно выше середины, но ниже верхней границы отпечатка переднего замыкателя и только в редких случаях на уровне верхней границы отпечатка.
<![if !vml]> <![endif]>
<![endif]>
Рис. 7.15. Изменчивость контура мантийной линии треугольной формы Syndosmya Аральского моря (по: Андреева, Андреев, 2001а). 1–2 — 1970 г., 3 — 1978 г., 4 — 1980 г., 5 — 1992 г., 6–8 —1994 г. Масштаб 1 мм.
<![if !vml]> <![endif]>
<![endif]>
Рис. 7.16. Изменчивость контура мантийной линии овальной формы Syndosmya Аральского моря (по: Андреева, Андреев, 2001а). 1 – 1970 г., 2 – 1978 г., 3 – 1980 г., 4 – 1981 г., 5 – 1990 г., 6 – 1992 г., 7 – 1994 г. Масштаб 1 мм.
Отпечатки мускулов-замыкателей у моллюсков всех форм в этот период имеют чрезвычайно изменчивые и извилистые контуры, вероятно, из-за развития дополнительных мышечных пучков. Границы отпечатков глубоко вдавлены во внутреннюю поверхность створки и часто окружены кораллово-жемчужными образованиями. Подобные образования имеются и внутри отпечатка.
В начале 1980-х годов контуры мантийной линии, в целом, сохраняют специфические особенности форм, но появляются дополнительные изгибы на восходящей и нисходящей ветвях, образующих примакушечный угол. Мускульные отпечатки с извилистыми границами значительно варьируют по размерам и форме и не имеют систематического значения.
Для 1989–1994 гг., в целом, сохраняется специфичность контура мантийной линии для удлиненно-овальной формы и овальной формы, но наблюдаются вариации размеров угла и изгибов ветвей мантийной линии, образующих его. Для треугольной формы в этот период времени типичный контур мантийной линии отмечается довольно редко, обычно наряду со слабо прослеживаемой истинной мантийной линией в передней части створки появляется четкий отпечаток, простирающийся по границе нижнего края пищеварительной железы. След истинной мантийной линии может исчезать совсем. Дополнительная мантийная линия в редких случаях может формировать второй угол в примакушечной части створки или прослеживаться повторной мантийной линией с еще одним примакушечным углом (см. рис. 7.15 6-8).
<![if !vml]> <![endif]>
<![endif]>
Рис. 7.17. Изменчивость отпечатка мускулов-замыкателей удлиненно-овальной формы Syndosmya Аральского моря (по: Андреева, Андреев, 2001а). 1 — 1970 г., 2 –4 — 1977–1980 гг., 5–6 — 1989 г. Закрашено – коралловые образования. Масштаб 1 мм.
Границы отпечатков замыкателей у моллюсков удлиненно-овальной формы и овальной формы в сборах 1989–1990 гг. еще крайне извилисты, у моллюсков треугольной формы происходит стабилизация границ. В 1992 г. стабилизация границ отпечатков характерна для всех моллюсков из Большого моря. В 1994 г. сглаживаются контуры отпечатков замыкателей и для моллюсков из Малого моря, хотя в более осолоненных районах (станции 15 и 16) процесс округления контуров отпечатков более замедлен.
<![if !vml]> <![endif]>
<![endif]>
Рис. 7.18. Изменчивость отпечатка мускулов-замыкателей треугольной формы Syndosmya Аральского моря (по: Андреева, Андреев, 2001а). 1 — 1970 г., 2 – 7 — 1977–1981 гг., 8–14 — 1989–1994 гг. Закрашено – коралловые образования. Масштаб 1 мм.
<![if !vml]> <![endif]>
<![endif]>
Рис. 7.19. Изменчивость отпечатка мускулов-замыкателей овальной формы Syndosmya Аральского моря (по: Андреева, Андреев, 2001а). 1 – 2 — 1970 г., 3 – 5 — 1977–1981 гг., 6–10 — 1989–1994 гг. Масштаб 1 мм.
Итак, в 1970-1994 гг. наблюдалось два периода изменчивости мантийной линии, совпадающих с пиками повышения изменчивости наружного лигамента: в первые годы акклиматизации моллюска и в начале 1990-х годов. И в том, и в другом случае наиболее вариабельны эти признаки у моллюсков, принадлежащих к треугольной форме. Максимальная же изменчивость границ контуров мускулов-замыкателей наблюдалась во второй половине 1970–1980-х годов, в период относительной стабильности и формоспецифичности мантийной линии и лигамента.
В дночерпательных пробах Syndosmya из Аральского моря за 1970–1994 гг. отмечены моллюски от 1.0 до 20.2 мм длиною. Максимальная длина моллюсков в 1970 г. была 14.6 мм, в 1977–1978 гг. — 20.2, в 1980 г. — 16.5, 1989 г. — 16.4, в 1994 г. — 14.8. Наибольшую длину имели моллюски овальной формы, наименьшую – треугольной формы (табл. 7.3).
Морфометрические индексы раковин Syndosmya довольно изменчивы (табл. 7.4). Наблюдаются различия среди форм Syndosmya по отношению высоты раковины к ее длине и выпуклости раковины к ее длине (табл. 7.5–7.7). Так, за весь период наблюдений за Syndosmya Аральского моря для удлиненно-овальной формы средние значения этих индексов колебались в пределах 0.70–0.74 и 0.32–0.35, для треугольной формы – 0.78–0.84 и 0.35–0.41, для овальной – 0.75–0.77 и 0.34–0.35 соответственно. В конце 1970-х годов наблюдались наиболее резкие различия форм Syndosmya по отношению высоты раковины к ее длине. Так, для удлиненно-овальной формы в 1977 и 1980 гг. – наблюдалось минимальное значение этого индекса, а для треугольной формы – максимальное значение этого индекса за весь период наблюдений. Для овальной формы отношение высоты раковины к ее длине довольно стабильно во времени.
Итак, наиболее четкие различия по морфометрическим индексам среди форм Syndosmya характерны для 1977–1978 гг. – времени завершения относительно стабильного периода существования Аральского моря, когда колебания его уровня и солености еще были в пределах вековых.
Таблица 7.3
Промеры раковин Syndosmya Аральского моря (в числителе – минимальное и максимальное значение признака, в знаменателе – среднее значение), мм (по: Андреева, Андреев, 2001а)
<![if !supportMisalignedColumns]><![endif]>
|
Год |
Длина раковины |
Высота раковины |
Выпуклость раковины |
n |
|
|
Удлиненно-овальная форма |
|||||
|
1970 |
9.6–14.6 |
7.0–10.2 |
3.1–4.9 |
28 |
|
|
1977 |
8.5–14.3 12.4±0.4 |
5.8–10.4 8.8±0.3 |
2.3–5.3 4.3±0.2 |
33 |
|
|
1980 |
7.8–16.5 12.6±0.8 |
5.1–11.3 8.9±0.6 |
2.3–5.7 4.2±0.3 |
24 |
|
|
1989 |
5.6–12.6 8.7±0.6 |
4.2–9.1 6.4±0.5 |
1.8–4.1 2.8±0.2 |
30 |
|
|
Треугольная форма |
|||||
|
1970 |
6.2–13.0 10.5±0.8 |
4.8–10.6 8.1±0.7 |
2.1–4.9 3.8±0.4 |
15 |
|
|
1977–1978 |
7.7–13.3 10.7±0.2 |
6.7–10.7 8.9±0.2 |
3.0–5.8 4.4±0.1 |
92 |
|
|
1980 |
6.2–10.4 8.5±0.7 |
5.2–8.5 7.1±0.5 |
2.7–4.3 3.3±0.3 |
14 |
|
|
1989 |
6.2–12.6 9.6±0.3 |
5.0–10.4 7.6±0.2 |
2.1–4.8 3.4±0.1 |
73 |
|
|
1994 |
8.6–14.8 11.5±046 |
6.3–11.8 9.0±0.3 |
2.9–5.8 4.4±0.2 |
50 |
|
|
Овальная форма |
|||||
|
1970 |
6.3–13.3 10.8±0.7 |
4.5–10.0 8.1±0.6 |
2.1–4.8 3.7±0.3 |
30 |
|
|
1977–1978 |
9.3–20.2 12.7±0.4 |
7.2.–14.7 9.8±0.3 |
3.1–7.8 4.6±0.2 |
97 |
|
|
1989 |
5.2–16.4 9.4±0.4 |
4.1.–12.8 7.2±0.3 |
1.7–6.2 3.2±0.2 |
103 |
|
|
1994 |
8.9–14.5 11.2±0.6 |
6.5–10.8 8.5±0.4 |
2.8–5.3 3.9±0.2 |
29 |
|
|
|
|
|
|
|
|
Примечание: n – число промеренных моллюсков.
При анализе сборов Syndosmya из Аральского моря за 1976–1978 гг. было обнаружено, что моллюски отличаются от типовой серии Syndosmya segmentum, описанной Я.И. Старобогатовым (1974), и образуют по контуру створок раковин три формы (Андреева, 1980), по внешнему облику сходные с видами Hypanis, обитавшими до середины 1970-х годов в Аральском море. Анализ сохранившихся сборов 1970 г., наиболее близких по времени к первому обнаружению (1967 г.) Syndosmya в Аральском море, показал, что уже и в этих сборах моллюски полиморфны. Варьирует контур створки, замок, лигамент, отпечатки мускулов-замыкателей (рис. 7.20). Изменчивость замка и лигамента настолько велика, что в качестве систематического признака их строение утрачивает свое значение. По контуру створок моллюски образуют почти непрерывный ряд от удлиненно-овального контура до треугольного, при этом особи из центральной и крайних частей ряда близки к формам, обитавших в Аральском море во второй половине 1970-х годов.
Ниже приведено описание эталонных материалов Syndosmya Аральского моря для 1977–1978 гг. – лет, завершающих период квазистационарного состояния моря, и когда уже наблюдалась стабилизация большинства морфологических признаков Syndosmya, а также для 1994 г. – года последних сборов Syndosmya в условиях резких изменений гидролого-гидрохимического режима моря.
<![if !vml]> <![endif]>
<![endif]>
7.1.6.1. Эталонные материалы удлиненно-овальной формы Syndosmya, станция 74 стандартной сетки станций, 1977 г.
Раковина тонкостенная, хрупкая, в виде вытянутого овала. Узкие, почти невыступающие макушки могут быть немного смещены к переднему краю раковины. Спинной край изогнут таким образом, что при переходе к переднему и заднему краям раковины углы не заметны (рис. 7.21). Величина апикального угла 140–145°.
Высота раковины составляет 0.69–0.70 ее длины, выпуклость – 0.34–0.36 ее длины и 0.49–0.52 ее высоты.
Наружный лигамент малосегментный, узкий с хорошо выраженным, крупным, округлым, передним сегментом.
Раковина почти симметричная, сильно зияющая спереди и сзади. На гладкой, просвечивающей, бесцветной или белой, ирризирующей поверхности раковины заметны концентрические линии нарастания. Узкая замочная площадка слабо изогнута так, что почти не образуется характерных для Syndosmya segmentum изгибов. На правой створке имеется два пластинчатых, маленьких кардинальных зуба, расположенных чаще всего под углом к нижнему краю замочной площадки и не доходящие примерно на треть до него.
<![if !vml]> <![endif]>
<![endif]>
Рис. 7.21. Удлиненно-овальная форма Syndosmya Аральского моря, станция 74, 1977 г. 1 – вид сбоку, 2 – вид сзади, 3 – вид сверху, 4 – отпечаток переднего мускула замыкателя, 5 – отпечаток заднего мускула замыкателя, 6 – лигамент, 7 – замок правой створки, 8 – замок левой створки. Масштаб 1 мм.
Первый передний латеральный зуб обычно рудиментарен, второй передний и задний латеральные зубы массивные, хорошо развиты. На левой створке – один кардинальный зуб треугольной формы и рудиментарные передний и задний латеральные зубы. Иногда их может не быть. Довольно широкая нимфа повторяет изгибы углубления внутреннего лигамента. Удлиненно-овальный, суженный в верхней части внутренний лигамент серповидно изогнут.
Мантийная линия характерной формы, образует тупой угол в примакушечной части створки, вершина которого расположена ниже верхней границы отпечатка переднего мускула-замыкателя. Углубленные отпечатки мускулов-замыкателей неправильной формы с довольно извилистыми границами могут быть окружены коралловыми образованиями. Передний отпечаток замыкателя уже заднего.
По внешнему облику раковина напоминает Syndosmya (=Abra) nitida milachevichi Nevesskaja, указываемую О.А.Скарлато и Я.И. Старобогатовым (1972) для Черного моря.
Раковина относительно толстостенная, треугольная или треугольно-овальная. Узкие, заостренные, выступающие макушки расположены почти посередине спинного края раковины. Спинной край изогнут таким образом, что при переходе к переднему и заднему краям образуются закругленные углы (рис. 7.22). Величина апикального угла 112–120°.
Высота раковины составляет 0.74–0.90 ее длины, выпуклость — 0.38–0.46 ее длины и 0.45–0.56 высоты раковины.
Наружный лигамент малосегментный, узкий, его передний сегмент обычно уже последующего.
Раковины почти симметричные, слабо зияющие спереди и сзади. На гладкой поверхности просвечивающей, белой или бесцветной, ирризирующей раковины хорошо прослеживаются концентрические линии нарастания.
<![if !vml]> <![endif]>
<![endif]>
Рис. 7.22. Треугольная форма Syndosmya Аральского моря, станция 63, 1978 г. Обозначение как на рис. 7.21.
Неширокая замочная площадка довольно круто изогнута, образуя в задней части почти прямой угол. На правой створке имеется два небольших, пластинчатых, кардинальных зуба, расположенных почти перпендикулярно нижнему краю замочной площадки. Передних латеральных зубов обычно два: первый очень тонкий и длинный, второй высокий и довольно массивный. Задний латеральный зуб хорошо выражен, его длина и форма несколько варьируют. На левой створке имеется один небольшой кардинальный зуб и обычно два узких тонких латеральных. Вместо заднего латерального может развиваться невысокий сильно сглаженный зубовидный выступ. Довольно широкая нимфа повторяет изгибы углубления внутреннего лигамента грушевидно изогнутой формы.
Мантийная линия характерной формы образует острый угол в примакушечной части створки, вершина которого расположена выше отпечатка переднего мускула-замыкателя. Отпечатки замыкателей неправильной формы с извилистыми границами, передний уже заднего. Изнутри створки раковины в примакушечной части сильно утолщены за счет кораллово-жемчужных образований в виде наплывов и наростов различной формы. В передней части створки часто наблюдается довольно мощный вырост-гребень, начинающийся на уровне границы
переднего замыкателя или чуть ниже и продолжающийся до примакушечного угла мантийной линии.
По внешнему облику моллюски напоминают Syndosmya segmentum [=Abra ovata (Phil.)] из Черного и Азовского морей, указываемую О.А. Скарлато и Я.И. Старобогатовым (1972).
Раковина тонкостенная, удлиненно-овальной формы. Узкие, слабо выступающие макушки смещены к переднему краю раковины. Спинной край изогнут таким образом, что при переходе к переднему и заднему краям образуются слабо заметные углы (рис. 7.23). Величина апикального угла 130–135°.
<![if !vml]> <![endif]>
<![endif]>
Рис. 7.23. Овальная форма Syndosmya Аральского моря, залив Чернышева, 1978 г. Обозначение как на рис. 7.21.
Высота раковины составляет 0.74–0.79 ее длины, выпуклость — 0.36–0.38 ее длины и 0.47–0.51 ее высоты.
Наружный лигамент малосегментный, удлиненно-овальной формы.
Раковина несимметричная, зияющая спереди и сзади. На гладкой, просвечивающей, бесцветной или белой, ирризирующей поверхности раковины заметны концентрические линии нарастания.
Читать продолжение седьмой главы