
Проблемы Эволюции |
О взрывном видообразовании, протекающем на наших глазах в высыхающем Аральском море. Омск: Изд-во Омского государственного педагогического университета, 2003. 382 с.
Особенности распределения одних и тех же или близкородственных видов, в частности, представителей «каспийской» фауны, при разной солености в Азовском, Каспийском и Аральском морях до сих пор не получили удовлетворительного объяснения и вызывают затруднения при рассмотрении эволюции моллюсков этих водоемов (Старобогатов, 1994; Ситникова, Старобогатов, 1999; и др.). При эволюционных построениях постулируется, что при вселении из пресных вод в соленые (замкнутые водоемы) происходит формирование новых видов, при этом механизм действия солености, как изолирующего фактора, не рассматривается (Логвиненко, 1968; Старобогатов, 1970, 1994; Старобогатов, Ситникова, 1992; и др.). Предваряя рассмотрение эволюции двустворок Арала, попытаемся выявить общие закономерности и особенности воздействия вод разной солености Азовского, Каспийского и Аральского морей на их фауну. При этом не лишним будет вспомнить слова Л.А. Зенкевича (1977) «… для каждого водоема потребуется своя собственная классификация, что определяется различиями в составе населения, во всей совокупности современных условий существования (абиотических и биотических) и в условиях геологического прошлого, когда формировалась солоноватоводная фауна» (стр. 273).
В морских гидробиологических работах неоднократно указывалось на соленость воды, как фактор, ограничивающий распространение той или иной фауны: пресноводной, солоноватоводной, морской и гипергалинной (Remane, 1934, 1940; Книпович, 1938; Dahl, 1956; Caspers, 1958; Хлебович, 1962, 1965; и др.). Наиболее подробно был исследован барьер между пресноводной и морской фаунами в водах океанического состава. Соленость 5–8 ‰ получила название «критической солености», и был выявлен универсальный характер барьера, разделяющего два главных типа фаун (Khlebovich, 1968, 1969; Хлебович, 1974). В последующем, для солености океанических вод 5–8 ‰, разделяющей фауны разного происхождения, был предложен O. Kинне (Kinne, 1971) термин «хорохалиникум» (horeo, греч. разделяю), а в русскоязычной литературе закрепился термин «хорогалинная зона» (Хлебович, 1986, 1989; Андреев, Андреева, 1990а).
Н.В. Аладиным (1988), на основании экспериментального изучения соленостных адаптаций Ostracoda и Branchiopoda и сведений об их распространении в природе, было выделено для морских вод пять барьерных соленостей 5–8, 16–20, 26–30, 36–40 и 50–55 ‰, не подтвержденных, за исключением первой, материалами по изменению физико-химических свойств воды в пределах барьеров.
В.В. Хлебович (1989) предложил классификацию морских вод, учитывающую хорогалинную зону (рис. 8.1). Нам (Андреева, Андреев, 2001б) представляется, что предложенная классификация учитывает не все барьеры, существующие при смешении речных и морских вод, в частности, барьер между пресными водами и олигогалинной зоной. Существование такого барьера вытекает из анализа работ по фауне Азовского моря. Так, одни авторы соленость 3–4 ‰ признавали основным ареалом солоноватоводной фауны (Зенкевич, 1959), другие – подчеркивали последовательный переход от чисто пресноводной фауны к фауне содержащей «каспийские» виды и далее к морской фауне (Марковский, 1953, 1954; Григорьев, 1965; Некрасова, 1976, 1977; и др.). Особый характер распределения каспийской фауны в Азово-Черноморском бассейне подчеркивается приводимой Ф.Д. Мордухай-Болтовским (1953, 1960) схемой галопатии (рис. 8.2).
В принимаемой Ф.Д. Мордухай-Болтовским (1960) классификации вод Азовского и Черного морей выделяются границы олигогалинной зоны (табл. 8.1), а Т.Г. Мороз и В.Л. Гильман (1988) разделяют солоноватые воды на относительные (0.5–2.5 ‰), промежуточные (2.5–7.5 с разбивкой на 2 подтипа: 2.5–5.0 и 5.0–7.5 ‰) и абсолютные (7.5–11.0 ‰).
По А.Ф. Карпевич (1960а), распространение пресноводных моллюсков в Азовском море ограничивалось соленостью 2 ‰. Примечательно, что при составлении «Определителя фауны Черного и Азовского морей» (1972) учитывались только виды, обитающие при солености более 2–3 ‰.
Учитывая приведенные выше данные, мы считаем (Андреева, Андреев, 2001б), что пресноводная (s. str.) фауна в Азово-Черноморском бассейне отделена от олигогалинной фауны, населяющей олигогалинные воды, барьерной соленостью 1–2 (3) ‰.
<![if !vml]>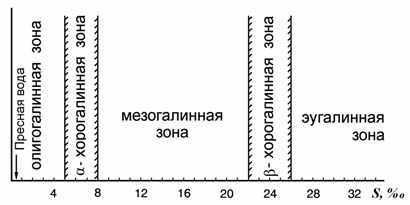 <![endif]>
<![endif]>
Рис. 8.1. α- и β-хорогалинные зоны в системе классификации вод по солености (по: Хлебович, 1989).
<![if !vml]>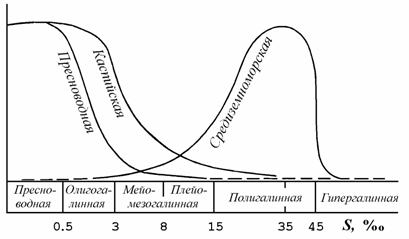 <![endif]>
<![endif]>
Рис. 8.2. Схема галопатии основных типов водной фауны Азово-Черноморского бассейна (по: Мордухай-Болтовской, 1953). По оси абсцисс – соленость, ‰; по оси ординат – число видов
Следует заметить, что как хорогалинная зона, так и другие барьерные солености выделяются с некоторой долей приближения, поскольку, как уже отмечалось (Андреев, Андреева, 1990а), точное положение этих зон выделить невозможно ни по распространению видов в природе, ни по экспериментальным данным. Наиболее четко положение хорогалинной зоны и, вероятно, других барьерных зон проявляется лишь при значительных изменениях экосистемы водоема в целом в процессе его осолонения.
Таблица 8.1
Схема подразделения солоноватых вод в Понто-Азовском бассейне (по: Мордухай-Болтовской, 1960)
|
Зона |
Подзона |
Соленость, ‰ |
|
Пресноводная |
– |
0–0.5 |
|
Олигогалинная, или слабосолоноватая |
– |
0.5–3(2) |
|
Мезогалинная, или собственно солоноватая |
Мейомезогалинная, или нижнесолоноватая |
3(2)–8(10) |
|
|
Плейомезогалинная, или верхнесолоноватая |
8(10) – около 15 |
|
Полигалинная, или сильно солоноватая |
– |
Около 15–30 |
Попытки разграничить воды Каспийского моря по градиенту солености и составу фауны предпринимались неоднократно. Так, Н.Л. Чугуновым (1922, цит. по: Бирштейн, 1945), считавшим соленость определяющим фактором в распределении донной фауны, выделялось для Северного Каспия три зоны:
1 – волжское пресноводное предустьевое пространство,
2 – солоноватоводная зона с двумя отделами,
3 – морская зона с постоянной, относительно высокой соленостью.
Хотя Я.А. Бирштейн (1945) большее значение в распределение донной фауны придавал характеру грунта и связанным с ним факторам – скорости течения и газовому режиму, выделенные им комплексы, населяющие грунты Северного Каспия, имели соленостные границы до 2–3 ‰, 3–7, 5–9, больше 9 и 10–12 ‰.
Обсуждая распределение фауны Каспийского моря по акватории, Л.А Зенкевич (1977) принял деление его вод на две зоны:
1. Меньше 7–8 ‰. Основной ареал солоноватоводной фауны, приуроченной к меньшей солености, со значительной примесью пресноводных форм.
2. От 7–8 до 13 ‰. Основной ареал солоноватоводной фауны, приуроченной к большей солености, с большей примесью морских иммигрантов.
Позднее Б.М. Логвиненко (1968) зону солености 3–7 ‰, характеризующуюся, по его мнению, в Каспии крайне скудным развитием жизни и, в частности, весьма бедным качественным и количественным составом донной фауны, предложил назвать «полипойкилогалинной зоной», которая по содержанию его работы соответствует хорогалинной зоне вод океанического состава. При этом было указано значение «полипойкилогалинной зоны» как барьера, защищающего вновь возникшие формы при вселении морских организмов в пресные воды.
В 1980-х годах положение хорогалинной зоны в Каспийском море рассматривал Н.В. Аладин (1983, 1989). Опираясь на работу А.Ф. Карпевич (1958), отмечавшую, что в ряду «океаническая вода®каспийская®аральская» наблюдаемое повышение толерантных соленостных диапазонов гидробионтов практически нивелируется по хлорности, он пересчитал первоначально по иону хлора (1983), а затем по иону натрия (1989) положение хорогалинной зоны вод с океаническим составом 5–8 ‰ для каспийских вод. По мнению автора, из-за различий в ионном составе хорогалинная зона в Каспии смещена до солености 7–11 ‰ и более широка, чем в водах океанического состава (рис. 8.3).
Помимо рассмотренных выше работ, во многих исследованиях донной фауны Каспийского моря содержится информация, позволяющая уточнить положение хорогалинной зоны и других барьерных соленостей в Каспийском море. Например, А.А. Шорыгин (1945), рассматривая бентос Северного Каспия, разбивает северо-каспийских двустворчатых моллюсков на три группы.
1. Группа видов, приуроченная к наименьшим соленостям: Adacna minima (=Hypanis vitrea glabra), Adacna plicata (=Hypanis plicata), Adacna laeviuscula (=Hypanis laeviuscula), Dreissena polymorpha. Средние солености от 3.68 до 6.18 ‰.
2. Группа видов, приуроченная в наибольшей степени к средним (с точки зрения Северного Каспия) соленостям: Monodacna edentula (=Hypanis angusticostata polymorpha), Monodacna caspia (=Hypanis caspia) и Didacna trigonoides. Средние солености от 8.5 до 10.5 ‰.
3. Группа видов, приуроченная к наивысшим соленостям Северного Каспия: Didacna barbotdemarnyi, Dreissena caspia, Mytilaster lineatus, Cardium edule (=Cerastoderma ? isthmicum). Средние солености от 11 до 12 ‰.
<![if !vml]>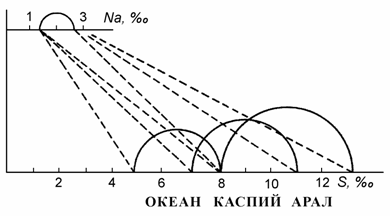 <![endif]>
<![endif]>
Рис. 8.3. Смещение критической солености в сторону высоких концентраций в водах Каспийского и Аральского морей по сравнению с водой океаническим составом солей (по: Аладин3, 1989).
По оси абсцисс: внизу – соленость, ‰; вверху –концентрация ионов натрия, ‰.
Показательно, что в 1935 г. максимальная биомасса Dreissena polymorpha приходилась на соленость 2 ‰ и от 6 до 10 ‰, что, вероятно, соответствует распределению в Северном Каспии ее двух подвидов: пресноводного D. p. polymorpha и солоноватоводного D. p. andrusovi. Для Adacna minima (=Hypanis vitrea glabra) — 2–4 ‰. Для наиболее соленолюбивых видов Didacna barbotdemarnyi, Dreissena caspia, Mytilaster lineatus, Cardium edule (=Cerastoderma ? isthmicum) и Monodacna caspia (=Hypanis caspia) максимум биомассы приходился на соленость 12–14 ‰. Интерпретация табличных данных А.А. Шорыгина (1945) по другим видам двустворок осложняется наличием у них подвидов,, выделенных только в последующем Б.М. Логвиненко и Я.И. Старобогатовым (1968), обитающих при различных соленостях.
Е.А. Яблонская и В.Ф. Осадчих (1973) указывают, что нижняя граница распространения средиземноморских и солелюбивых реликтовых моллюсков [церастодермы, митилястера, абры (=синдосмии), дидакны] приурочена к водам соленостью 8–9 ‰ и более. При этом отмечалось, что морские организмы – выходцы и вселенцы из Средиземноморского бассейна наиболее высокую биомассу образуют обычно в западной половине Северного Каспия, где наблюдается постоянный приток среднекаспийских соленых вод и имеется прямой путь для иммиграции личиночных форм из Среднего Каспия, в котором эти организмы имеют широкий ареал и высокую численность.
Л.Г. Виноградов (1959, 1963; цит. по Яблонская, Осадчих, 1973) отмечал, что организмы солоноватого комплекса (моллюски монодакна, дрейссена, дидакна), населяющие районы, несколько удаленные от устьев рек, не переносят ни сильного опреснения, ни сильного осолонения. Поэтому в годы сокращения волжского стока и повышения солености Северного Каспия биомасса этих видов уменьшается. Интересны и подразделения биоценозов Северного Каспия, предложенные Л.Г. Виноградовым (1959):
1 – пресноводные [пресноводные моллюски, речная дрейссена (=Dreissena polymorpha polymorpha), олигохеты, личинки хирономид];
2 – прибрежные и слабосолоноватоводные [адакна (=Hypanis vitrea glabra), уралокаспийская форма дрейссены (=Dreissena polymorpha ? polymorpha), высшие ракообразные, олигохеты, амфаретиды, личинки хирономид];
3 – солоноватоводные [монодакна (=Hypanis caspia caspia и H. angusticostata), трехгранная дидакна (=Didacna trigonoides), морская форма дрейссены (= Dreissena polymorpha andrusovi)];
4 – морские (средиземноморские митиластер, церастодерма, нереис);
5 – соленолюбивые реликтовые [морская дидакна (=Didacna longipes, Didacna protracta и Didacna barbotdemarnyi), каспийская дрейссена (=Dreissena caspia)].
Очень показательна схема распределения кардиид по акватории Каспия, составленная А.Ф. Карпевич (1975) по материалам Я.И. Бирштейна (рис. 8.4), из которой следует, что распределение кардиид приурочено к определенным зонам солености: от чуть менее 5 до 10 ‰ – Hypanis minima (=Hypanis vitrea glabra), 10 ‰ и несколько ниже – Monodacna edentula (=Hypanis angusicostata polymorpha) и Didacna trigonoides, в основном 10 ‰ и выше – Didacna barbotdemarnyi и Cerastoderma lamarcki (=Cerastoderma ? isthmicum).
Вселившийся средиземноморский двустворчатый моллюск Mytilaster lineatus также преимущественно был распространен при солености не ниже 10 ‰ и положение границ его ареала в Северном Каспии зависело от солености (рис. 8.5).
Учитывая распространение чисто пресноводных видов, ограниченное преимущественно изогалиной 2 ‰, можно предположить, что в Каспийском море помимо хорогалинной зоны, разделяющей эвригалинные виды морского происхождения и олигогалинные виды, существует и барьерная соленость, отделяющая олигогалинную фауну от пресноводной. Как отмечалось выше, точное положение барьерных зон выделить практически невозможно и поэтому берется достаточно широкий диапазон. В Каспийском море хорогалинная зона располагается в области 8–10 ‰, а первая барьерная соленость отделяющая пресноводную фауну (s. str.) от олигогалинной — 2–4 ‰ (Андреева, Андреев, 2001б).
Проведенный на основе литературных данных (Сидоров, 1929; Бенинг, 1934; Деньгина, 1954, 1957, 1959а, б; Хусаинова, 1958, 1960; Яблонская, 1960; и др.) анализ (Андреев, Андреева, 1990а) распределения донной фауны в зависимости от солености для периода, когда условия обитания в Аральском море были близки к среднемноголетним, показал, что в собственно море при средней солености около 10 ‰ массовое развитие имели небольшое число видов: Dreissena obtusecarinata, моллюски рода Hypanis (H. vitrea bergi, H. minima minima, H. sidorovi), Chironomus behningi. Доминирующее положение этих видов сохранялось и в опресненных заливах до солености 4–5 ‰, но в култуках, при солености, превышающей соленость открытого моря всего лишь на 2 ‰ (больше 12 ‰), из вышеназванных видов в заметном количестве отмечалась только Dreissena obtusecarinata, да и то до солености не более 16 ‰. При солености более 12 ‰ в култуках получали условия для массового развития Cerastoderma isthmicum, моллюски рода Caspiohydrobia, Theodoxus pallasi, Dreissena caspia pallasi, последняя в узком соленостном диапазоне 12–20 ‰.
<![if !vml]> <![endif]>
<![endif]>
Рис. 8.4. Распределение кардиид в Северном Каспии в зависимости от солености в 1935 г. (по материалам Я.И. Бирштейна, из: Карпевич, 1975). 1 – Hypanis minima; 2 – Monodacna edentula; 3 – Didacna trigonoides; 4 – D. barbotdemarnyi; 5 – Cerastoderma lamarcki; 6 – изогалины, ‰.
Скачкообразное изменение фауны в култуках Аральского моря, приуроченное к солености воды 12–16, 23–25 и 32 ‰, было отмечено еще Н.З. Хусаиновой (1958). К сожалению ею не были приведены фактические материалы, подтверждающие обоснованность выделенных диапазонов солености, влияющих на качественный состав фауны.
<![if !vml]>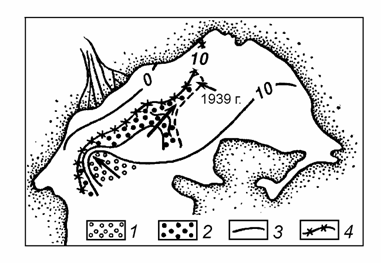 <![endif]>
<![endif]>
Рис. 8.5. Изменение ареала распространения Mytilaster lineatus. с 1932 по 1939 гг. в зависимости от солености (по: Карпевич, 1975).
1 – 1932 г., 2 – 1939 г. 3 – изогалина 10 ‰ в 1932 г., 4 – изогалина 10 ‰ в 1939 г.
Как показали более поздние исследования (Андреев, 1981; Андреев, Андреева, 1981, 1990а, 1991б), первый из указанных Н.З. Хусаиновой (1958) диапазонов солености аральской воды, но более узкий — 12–14 ‰, разделяет пресноводную и морскую фауны, соответствуя хорогалинной зоне 5–8 ‰ для вод с океаническом составом солей. Получил подтверждение и второй диапазон соленостей, указанный Н.З. Хусаиновой (1958). Всего для Аральского моря нами (Андреев, Андреева, 1998а, Андреев, 1999) было выделено 4 барьерные солености: 3–5, 12–14, 24–28 и 40–45 ‰, названных позднее олигогалинным, мезогалинным, эугалинным и гипергалинным барьерами (Андреева, Андреев, 2001б).
Н.В. Аладиным (1990), на базе концепции относительности и множественности зон барьерных соленостей (Аладин, 1988), определено в Аральском море 5 барьерных соленостей: 7–11, 22–28, 36–42, 50–56, 66–77 ‰. Автором (Аладин, 1990) был сделан формальный пересчет концентрации ионов натрия пяти барьерных соленостей морских вод в соответствующую соленость аральских вод и, поскольку, по его мнению, «...именно гидробионты с их осморегуляторными способностями привносят в водные экосистемы эффекты барьерных соленостей» (с. 11), подробно обсуждать выделенные барьеры не имеет смысла. Остановимся лишь на хорогалинной зоне. Получив пересчетом ее значение для вод Аральского моря (см. рис. 8.3) равным 7–11(?) ‰ (Аладин, 1990) или 8–13 ‰ (Аладин, 1983, 1986, 1996) автор считал, что по сравнению с океаническими водами в аральской воде интервал хорогалинной зоны увеличивается с 3 до 5 ‰, не приводя серьезных аргументов в пользу такого увеличения. В этом случае интервал хорогалинной зоны (критической солености биологических процессов) захватывает период существования экосистемы моря при его квазистационарном режиме Арала в первой половине 20 века, когда наблюдалось процветание фауны, вплоть до размножения почти всех пресноводных рыб при солености открытого моря (средняя 10.2 ‰). Кризисное же состояние фауны моря документально зарегистрировано (см. глава 3) при приближении вод Арала к солености 12–14 ‰, а повышенная изменчивость эвригалинных двустворок отмечена после перехода экосистемой моря барьера 12–14 ‰ (см. глава 6–7).
Исследованиями последних лет в области гидрологии и гидрохимии Аральского моря (Гидрометеорология …, 1990; Цыцарин, Лобов, 1993а, б, 1994, 1995; Лобов, 1994; Лобов с соавт., 1995) выявлено, что в зонах барьерных соленостей изменяются физико-химические свойства его вод.
Олигогалинный барьер 3–5 ‰ ограничивает распространение собственно пресноводной (s. str.) фауны. Наличие этой барьерной солености и позволило иметь своеобразный, резко отличающийся видовой состав беспозвоночных животных, населяющих Арал.
По данным А.Г. Цыцарина и А.Л. Лобова (1995), в Аральском море барьер между речными и морскими водами в среднем расположен в области значений 2.5 ‰, а его нижняя и верхняя границы соответственно в области 1.5 и 3–4 ‰. В переходной зоне отношение содержания иона кальция и карбонат иона к иону хлора снижается на порядок. Изменение положение границы барьера от 3 (весна) до 2 (лето) и 4 ‰ (осень) обусловлено существенным влиянием температуры воды на растворимость карбоната кальция, а также зависит от содержания карбонатов в речной воде.
Мезогалинный барьер 12–14 ‰ в Аральском море соответствует хорогалинной зоне 5–8 ‰ морских вод с океаническим составом (Андреева, Андреев, 2001б).
В пределах этого барьера начинается метаморфизация вод Арала. Аральская вода была насыщена карбонатом кальция еще при квазистационарном режиме моря при солености 10 ‰ (Блинов, 1956). Небольшое повышение солености воды в 1960-х годах вызвало нарушение равновесия солевой системы моря и началось значительное осаждение солей. Считается, что при концентрации морской воды в период с 1961 по 1970 гг. выпало в осадок 2.39 млн. т карбонатов, а в 1971–1980 гг. — 4.34 млн. т (Гидрометеорология…, 1990). Рост солености в этот период сопровождался ростом абсолютного содержания всех ионов, при этом при солености около 14 ‰ сильно возрастала дисперсия содержания Mg2+ и Ca2+ (Лобов с соавт., 1995).
Эугалинный барьер 24–28 ‰ (в ранних работах 25–30 ‰) является верхней границей распространения солоноватоводных видов.
На соленость 24.7 ‰ приходится переход вод Аральского моря из класса солоноватых в класс морских, когда температура наибольшей плотности становится равной температуре замерзания. При этой солености происходит принципиальная перестройка зимнего охлаждения водоема от пресного типа к морскому (Цыцарин, Лобов, 1995). Именно в пределах этой барьерной солености наблюдались существенные перестройки солевой системы моря. Так, во второй половине 1980-х годов для солевого состава воды Аральского моря был важен процесс выпадения в осадок CaSO4, так как он привел к уменьшению концентраций Ca2+ и SO42– и метаморфизации солевого состава. Перестройка солевой системы моря в 1989 г. (средняя соленость примерно 27 ‰) сопровождалась изменением коэффициентов в соотношении хлорности и концентрации HCО3–, хлорности и концентрации Ca2+ (Лобов, 1994).
Гипергалинный барьер 40–45 ‰ ограничивает распространение видов морского происхождения и является нижней границей распространения для гипергалинных видов.
При солености 40–45 ‰ солевая система Аральского моря в очередной раз переходит в неустойчивое состояние (Цыцарин, Лобов, 1995).
Множественный характер барьеров позволяет предполагать о разграничении ими водной среды обитания на отдельные водные массы со специфическими физико-химическими свойствами (Андреев, Андреева, 1990а, 1998а). Если в пределах барьерных соленостей происходит резкое качественное изменение физико-химических свойств вод моря, то между барьерными соленостями заключены водные массы, характеризующиеся относительной однородностью физико-химических свойств. В пределах водной массы происходит количественное изменение параметров, например, солености, подготавливающее качественный скачок в пределах барьера. Каждую водную массу населяют специфические флора и фауна в соответствии с осморегуляторными возможностями составляющих их видов.
На базе разработанной классификации вод Аральского моря (Андреев, Андреева, 1998а) представляется возможным предложить единую классификацию вод океанического состава, Аральского и Каспийского морей с поправкой на положение барьерных соленостей (рис. 8.6). Помимо указанных водных масс: пресноводной, олигогалинной, мезогалинной и эугалинной, существуют еще гипергалинные и ультрагалинные водные массы и соответственно гипергалинный и ультрагалинный барьеры, но, поскольку по их положению мы не располагаем достаточной информацией, в данной работе они не рассматриваются. Предлагается присвоить барьерным соленостям привычные названия в соответствии с отделяемыми ими водными массами: олигогалинный, мезогалинный, эугалинный, гипергалинный и ультрагалинный барьеры.
Гидрохимические исследования достаточно ясно показывают, что при смешении речных и морских вод в эстуарных зонах возникают барьеры, обусловленные изменением физико-химических свойств воды. Так, удаление главной массы взвешенного материала наблюдается при солености около 2 ‰. К тому же, в речной воде соли, в основном, находятся в виде взвесей, в морской воде при солености более 5 ‰ соли на 98 % – в растворенном состоянии (Гордеев, 1983). При смешении вод в эстуарных зонах прослеживаются все переходные стадии между гидрокарбонатно-кальциевыми и хлоридно-натриевыми водами. Особо резкие изменения относительно содержания основных ионов происходят при солености менее 2 ‰. Преобразование ионного состава вод продолжается до солености 5 ‰, при солености выше 10 ‰ относительное содержание ионов практически не меняется (Цыцарин, Лобов, 1995). Исследования А.К. Виноградова (1986а, б) обнаружили преобразования физико-химических свойств воды и растворов хлористого натрия также в диапазоне солености 22–26 ‰.
<![if !vml]>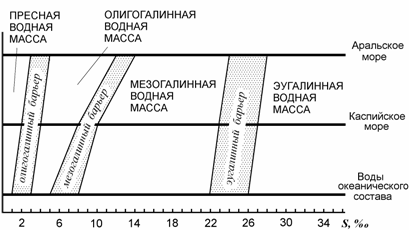 <![endif]>
<![endif]>
Рис. 8.6. Классификация вод с учетом барьерных соленостей (по: Андреева, Андреев, 2001б). По оси абсцисс– соленость, ‰;
В процессе изменения характера вод суши на морской возникают карбонатный и сульфатный физико-химические барьеры, определяемые достижением предела растворимости солей и имеющие общегеографический характер. Однако, этот процесс в различных морях не полностью тождествен и имеет региональные особенности (Питьева, 1978; Никаноров, Посохов, 1985; Цыцарин, Лобов, 1995).
Исследуя положение физико-химического барьера река – море для Аральского, Азовского морей и северной части Каспийского, А.Г. Цыцарин и А.Л. Лобов (1995) указывают, что для этих объектов нижняя граница переходной зоны расположена в области значений 1.5 ‰ для кальция и 1 ‰ для гидрокарбонатов. Ширина переходной зоны определяется относительным содержанием рассматриваемых ионов в речной и морской воде – чем меньше их значения в реке и больше в море, тем шире переходная зона. Сама зона представляет собой переход от уже метаморфизированных речных вод к морским водам, а с верхней границей переходной зоны совпадает нижняя граница диапазона применимости гипотезы квазипостоянства солевого состава морской воды. Наиболее резко барьер между речными и морскими водами выражен в Таганрогском заливе Азовского моря, где он расположен в области значений 2.0–2.5 ‰ суммы солей. Положение барьера для Северного Каспия составляет 2–4 ‰ суммы солей, а для Арала – от 1.5 до 3–4 ‰.
Универсальным барьером, разделяющим морскую и пресноводную фауны, является зона критической солености, или хорогалинная зона 5–8 ‰ для вод с океаническим составом солей (Хлебович, 1965, 1974; Khlebovich, 1968, 1969, 1990), «...определившая направление развития органического мира в далеком прошлом и ряд существенных свойств современных биохимических и клеточных структур, видовых адаптаций и структуры фаунистических комплексов» (Хлебович, 1974, с. 197). Недавно были вторично описаны процессы, характеризующие воды в области зоны критической солености под названием «маргинальный фильтр океана» (Лисицын, 1994, цит. по Хлебович, 1996).
Как уже указывалось выше, в пределах барьерных соленостей происходит резкое качественное изменение физико-химических свойств вод моря. Между барьерными соленостями заключены водные массы, характеризующиеся относительной однородностью физико-химических свойств, в то же время в пределах водной массы происходит количественное изменение параметров, например солености, подготавливающее качественный скачок в пределах барьера. Каждую водную массу, которая является специфической ареной жизни (специфической адаптивной зоной), населяют специфические флора и фауна в соответствии с осморегуляторными возможностями составляющих их видов. Населяющие водные массы флора и фауна представляют собой интегральный показатель взаимодействия комплекса абиотических и биотических факторов вдоль оси смешения пресных вод реки, осолоненных вод моря и, там где они есть, высокоминерализованных вод заливов, маршей.
Барьерные солености разделяют определенные адаптивные зоны (=арены жизни), для освоения которых живому необходимо преодолеть ограничивающие их барьеры. Преодоление барьерных соленостей в процессе эволюции жизни в водной среде осуществлялось за счет приобретения и совершенствования разнообразных осморегуляторных механизмов, которые, вероятно, следует рассматривать как морфофизиологические адаптации арогенного ранга. Разнообразие морфофункциональных адаптаций обеспечивало гидробионтам освоение всего комплекса природных поверхностных вод. В зависимости от особенностей развития морфофизиологических адаптаций как у групп организмов, так и у отдельных видов осваивались соответствующие водные массы или же при совершенных осморегуляторных механизмах – несколько водных масс.
Особое значение барьерные солености должны были иметь при формировании солоноватоводных эндемичных фаун. Ярким подтверждением этого положения служат специфические фауны Аральского и Каспийского морей, а также распространение каспийской фауны в Азово-Черноморском бассейне.
При разработке концепции критической солености В.В. Хлебович (1974) предложил гипотезу о происхождении в далеком прошлом основных форм метаболизма при солености выше критической, но близкой к ней. Основные формы осморегуляции, по-видимому, вырабатывались при переходе через барьер критической солености на пути в ту или иную сторону (рис. 8.7).
Выработка, на базе истинных пойкилоосмотических организмов, гипертонической регуляции позволила многим большим группам животных перейти мезогалинный барьер (=барьер критической солености) и колонизовать пресные воды. Выработка механизмов гиперосмотической регуляции, прежде всего аппаратов активной сорбции Na+ из среды и реабсорбции его при образовании гипотонической мочи, является одним из центральных моментов эволюции животных. Переход через критическую соленость настолько связан с коренным преобразованием функций и необходимостью выработки принципиально новых органов, что можно представить, что он происходил под влиянием могучего пресса, вероятно, прежде всего биотических отношений (Хлебович, 1974).
<![if !vml]>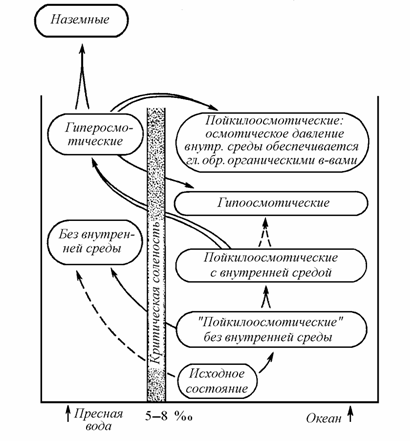 <![endif]>
<![endif]>
Рис. 8.7. Основные пути эволюции соленостных отношений животных (по: Хлебович, 1974).
Принимая схему эволюции животных В.В. Хлебовича (см. рис. 8.7), обсудим более подробно освоение гидросферы Mollusca с учетом всех барьерных соленостей. Рассматриваемая группа относится к настоящим пойкилосмотическим организмам с внутренней средой – целомической жидкостью, кровью и пр. При проникновении через барьер критической солености в пресные воды моллюски должны переходить к гиперосмотической регуляции. После преодоления барьера критической солености формируются две обширные группы моллюсков: пресноводные и морские, эволюция которых далее протекала независимо.
Из выделенных водных масс в силу геологических и исторических причин наибольшим постоянством и длительностью существования обладают пресные и эугалинные водные массы. Напротив, олигогалинные и мезогалинные водные массы в геологическом масштабе времени не постоянны и неоднократно могли возникать и исчезать в каком-либо регионе. В связи с чем, наибольшего разнообразия моллюски достигли в пресных и эугалинных водных массах, и малакофауна этих водных масс является донором для формирования фаун олигогалинных и мезогалинных водных масс.
Освоение (проникновение в) соседней водной массы могло идти двумя путями. Во-первых, путем выработки эвригалинности, то есть приобретения морфофизиологических адаптаций, позволяющих существовать в условиях широкого соленостного спектра (двух и более водных массах), что особенно важно в условиях непостоянства среды обитания. Во-вторых, после проникновения в соседнюю водную массу в условиях ее постоянства в геологических масштабах времени был возможен путь приобретения стеногалинности или выработки адаптаций для обитания в пределах конкретной водной массы с формированием специфической олигогалинной или мезогалинной фауны, не встречающейся в других водных массах. Присутствие специфической, большей частью эндемичной, олигогалинной или мезогалинной фауны – это свидетельство особых путей развития экосистемы водоема.
Для моллюсков в силу специфики организации все барьерные солености являются трудно преодолимыми, и поэтому их видовой состав резко различается по ту и другую сторону барьерной солености, тогда как пресноводные рыбы свободно преодолевают олигогалинный
барьер.
В проявлении феномена барьерных соленостей необходимо учитывать историю формирования фаун. Так, в опытных работах по соленостной резистентности (аральская вода) эвригалинных беспозвоночных разного происхождения (Андреев, Андреева, 1983, 1990б) было прослежено резкое снижение выживаемости в пределах мезогалинного барьера (= хорогалинной зоны) средиземноморско-атлантических видов на клеточном, тканевом и организменном уровне, в то время как эвригалинные обитатели материковых вод не имели достоверного снижения выживаемости в этом диапазоне (рис. 8.8). Более того, в условиях, когда две водные массы имеют достаточно большой объем и существуют в пределах одного водоема длительное в геологических масштабах время и с постепенным переходом одной в другую, то у обитателей такого водоема вырабатываются морфофизиологические адаптации для свободного преодоления барьерных соленостей.
Наиболее показательны в этом плане представители семейства Podonidae Каспийского моря, выработавшие в процессе эволюции способность к амфиосмотической регуляции гемолимфы, которая позволяет им легко преодолевать мезогалинный барьер. Представители этого семейства в водах океанического состава способны только к гипоосмотической регуляции (Хлебович, Аладин, 1976; Potts, Durning, 1980; Аладин, 1982а, б). При проникновении в Каспий Pleopis polyphemoides (Leuck.) менее чем за двадцать лет восстановил амфиосмотическую регуляцию гемолимфы, хотя до этого при обитании в открытых морях демонстрировал как в природе, так и в эксперименте, способность только к гипоосмотической регуляции (Аладин, 1982б, 1986).
При впадении рек в океан переход пресных водных масс в эугалинные обычно совершается быстро, формирующиеся при этом мезогалинные и, особенно, олигогалинные водные массы непостоянны во времени и в пространстве, в связи с чем специфическая мезогалинная и, особенно, олигогалинная фауны либо не развиваются, либо представлены ограниченным числом видов. Олигогалинные и мезогалинные водные массы в этих случаях заселяются эвригалинными видами морского или пресноводного происхождения. Только во внутренних морях: Балтийском, Черном, Азовском или эстуариях, где возникают устойчивые водные массы разной солености, формируются, в той или иной мере, специфические мезогалинная и олигогалинная фауны, впрочем, для олигогалинной фауны и тут условия обитания малоподходящие, в силу чего она представлена небольшим числом видов.
<![if !vml]>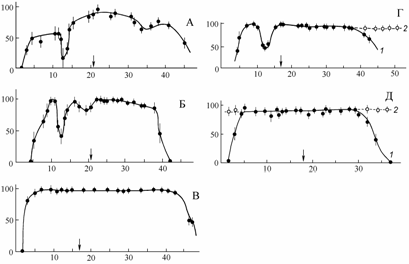 <![endif]>
<![endif]>
Рис. 8.8. Выживаемость взрослых особей Nereis diversicolor (А), Cerastoderma istmicum (Б), Caspiohydrobia sp. (В), Syndosmya segmentum (Г) и Theodoxus pallasi (Д) в аральской воде различной солености (по: Андреев, Андреева, 1990б).
Стрелка – исходная соленость из мест обитания животных. По оси абсцисс – соленость, ‰; по оси ординат – количество выживших животных на 15 сутки опыта, процент от общего числа: 1 – при прямом переносе, 2 – при ступенчатой акклимации.
Другая картина складывается в континентальных водоемах, не имеющих связи с океаном. Режим этих водоемов обусловлен объемом стока, долговременными климатическими и эрозионными процессами и т. п. Даже самые крупные водоемы в своей истории не только неоднократно испытывали периоды осолонения и опреснения с переходом через соленостные барьеры, но и периоды превращения в гипергалинные водоемы или периоды полного осушения. Тем не менее, только во внутренних водоемах, лишенных связи с океаном, возможно существование олигогалинных и мезогалинных водных масс длительное время, достаточное для формирования специфической фауны.
Особенностью освоения континентальных водоемов, облегчающей преодоление барьерных соленостей, следует считать однонаправленный процесс вселения из пресных водных масс в олигогалинные, а затем и в мезогалинные водные массы. Вероятное отсутствие конкуренции в первый момент внедрения в следующую водную массу значительно облегчало преодоление барьерных соленостей. С другой стороны, отчленившиеся водоемы с типичной морской фауной в процессе опреснения могли сохранить только небольшое количество видов, способных обитать в олигогалинной или мезогалинной водной массе, на основе которых происходило формирование эндемичных родов и семейств.
Рассмотрим с позиций барьерных соленостей историю формирования фауны моллюсков Азовского, Каспийского и Аральского морей.
Истории формирования фауны водоемов Паратетиса посвящена обширная литература, но до сих пор нет единства мнений (см. Старобогатов, 1970, 1994; Григорьев, Гожик, 1976; Невесская с соавт., 1986; и др.). Не вдаваясь в подробности геологической истории водоемов и смены фаун, отметим лишь, что примерно 15 млн. лет назад сформировалось Нижнесарматское море, протянувшееся от Среднедунайской низменности до Приаралья. Оно имело несколько пониженную, по сравнению с океаном соленость, но было населено нормальной морской фауной. Позже соленость моря постепенно падает, и в Верхнесарматском море, и в обособившемся Паннонском озере-море формируются эндемичные фауны (Старобогатов, 1994). Изолированное от океана развитие фаун солоноватых морей шло более 10 млн. лет.
В четвертичное время соленость воды в Каспийском бассейне не превышала, по мнению Я.И. Старобогатова (1994), солености 12 ‰ (рис. 8.9). В Черноморском бассейне изолированное развитие фауны дважды прерывалось вторжением морских вод. Первый раз около 80 тыс. лет назад с образованием Карагантского моря и второй раз примерно 11–10 тыс. лет назад с образованием современного моря.
<![if !vml]>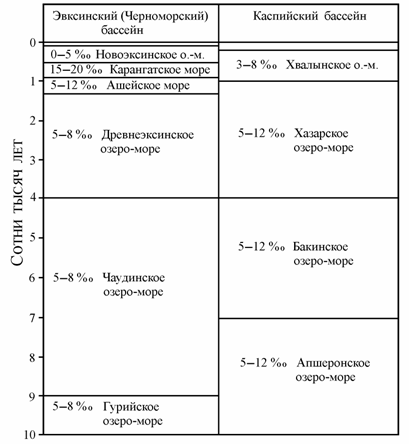 <![endif]>
<![endif]>
Рис. 8.9. Схема синхронизации водоемов Эвксинской (=Черноморской) и Каспийской котловин в четвертичное время (по: Старобогатов, 1994).
С позиции предлагаемой концепции водных масс при вторжении морских вод мезогалинная специфическая фауна Ашейского, а в последующем и Новоэксинского морей вымирала; олигогалинная фауна имела возможность отступать в эстуарии и устья рек, где частично сохраняется и в настоящее время. Уцелевшие представители понто-каспийской фауны в современных условиях Азовского моря уже не являются олигогалинной (s. str.) фауной, поскольку они должны были приспособиться к переживанию более или менее длительного времени в пресных водных массах, приобретя некоторые черты эвригалинности. Тем не менее, представители понто-каспийской фауны обычно занимают промежуточное положение между пресноводной (s. str.) и морской фаунами и имеют массовое развитие в олигогалинных водных массах (Марковский, 1953; Мордухай-Болтовской, 1960; Григорьев, 1965).
Считается, что род Dreissena сформировался в пресных водах восточного Средиземноморья не позже позднего миоцена, откуда его представители распространились по речным системам Эгейского бассейна и затем проникли в Паннонский бассейн, где вселились в солоноватые воды (Старобогатов, 1994). При этом дрейссены должны были последовательно осваивать пресные, олигогалинные и мезогалинные водные массы, преодолевая олигогалинный и мезогалинный барьеры. Барьерные солености при этом выступали как надежный изолирующий фактор в распределении видов по градиенту солености. Например, в Каспийском море в пресных водных массах обитает Dreissena polymorpha polymorpha, в олигогалинных – D. andrusovi, в мезогалинных в пределах глубин до 35 м, расходясь по грунтам – D. caspia caspia и D. elata, на глубинах от 20 до 300 м – D. rostriformis. В Аральском море этот ряд представляют D. p. aralensis – D. obtusecarinata – D. caspia pallasi. При преодолении первой барьерной солености у дрейссен, обладающих плавающей личинкой, длительное время находящейся в водной толще, идет процесс видообразования, но поскольку по ту и другую сторону барьера образ жизни сохраняется практически неизменным, то в строении раковин больших отличий и не наблюдается, изменения затрагивают главным образом только мягкие ткани. Точно ответить на этот вопрос смогут только специальные морфологические и генетические исследования.
В Азово-Черноморском бассейне в связи с вторжением соленых океанских вод и населяющей их фауны все мезогалинные специфические Dreissena вымерли, а уцелевшие представители рода обитают в олигогалинных и пресных водных массах.
Обособление подрода Pontodreissena связано с освоением водных пространств Паннонского бассейна при переходе с твердых субстратов на мягкие с рассеянными твердыми включениями. В отличие от подрода Dreissena (s. str.), центральный подвид которого D. polymorpha polymorpha на протяжении голоценовой истории мог сохраняться в пресных водах, где условия мало менялись, виды подрода Pontodreissena могли сохраняться только в гигантских озерах (Старобогатов, 1994).
Очевидно, представители Pontodreissena первоначально освоили олигогалинные и мезогалинные водные массы. Лишь в Эвксинском бассейне, где в связи с вторжением морской фауны сохранился единственный представитель D. bugensis, который, приобретя черты эвригалинности, осваивает и пресные водные массы.
Регистрируемое в настоящее время проникновение Dreissena bugensis в пресные воды (Старобогатов, 1994) является повторным возвращением Dreissena в пресные водные массы и должно сопровождаться видообразовательным процессом, если олигогалинные водные массы в Азово-Черноморском бассейне будут более устойчивы во времени и пространстве.
В Каспийском море, где с акчагыльского времени отмечались колебания уровня от –30 до +70 м, по сравнению с современным (Старобогатов, Ситникова, 1992), напротив, сохранились представители мезогалинной специфической фауны: D. r. grimmi, D. r. pontocaspica, D. r. compressa и D. r. distincta. Вероятно, в Каспии помимо колебаний солености большое значение имели и конкурентные взаимоотношения с представителями подрода Dreissena (s. str.), с которыми сохранившиеся представители викарируют по вертикали, встречаясь с глубины 20 м, тогда как Dreissena (s. str.) заселяют мелководья.
Хотя D. bugensis обитает в Черноморском бассейне в пресной и олигогалинной водных массах при солености 5 ‰ и ниже, а подвиды D. rostriformis в Каспийском море в мезогалинных водных массах при солености более 10 ‰, с позиций концепции водных масс и учета специфики ионного состава вод морей, представители подрода Pontodreissena выстраиваются по градиенту солености в один непрерывный ряд, с разделением по барьерной солености.
Анализ распределения современных солоноватоводных кардиид южных морей обнаруживает существование двух обширных групп, приуроченных к олигогалинным (род Hypanis частью) и мезогалинным (род Didacna и род Hypanis частью) водным массам. При этом одновременное обитание в водоеме всех представителей обоих родов возможно только при наличии обширных олигогалинной и мезогалинной водных масс, что имеет место в современном Каспии и, вероятно, было характерно для палеоводоемов. В прежние геологические эпохи эти моллюски были распространены гораздо шире и считаются производными морской фауны, освоившими солоноватые (мезогалинные и олигогалинные) воды в неогене, в частности, Didacna известны с понтийского времени, Hypanis – с апшеронского (Колесников, 1939; Старобогатов, 1970; Мерклин, 1974; Григорьев, Гожик, 1976; и др.). В современном Азово-Черноморском бассейне распространение моллюсков рода Hypanis приурочено к олигогалинным водным массам лиманов, приустьевых районов и солоноватых озер. Чрезвычайно интересным фактом с позиции концепции водных масс является отмечаемое в последнее время расселение в пресные воды Hypanis colorata.
Вымершая в 1970-е годы фауна Hypanis Аральского моря обитала только в олигогалинных водных массах, занимавших в первой половине двадцатого века почти всю его акваторию.
Более интересно распространение Hypanis и Didacna в современном Каспии, большую часть акватории которого занимают мезогалинные водные массы, сменяющиеся в Северном Каспии олигогалинными. Практически все Didacna приурочены к мезогалинным водным массам, только Didacna trigonoides проникает в олигогалинные районы. Среди эндемичных гипанисов выстраивается ряд видов, сменяющих друг друга вдоль градиента солености: в мезогалинных водных массах это, например, Hypanis caspia и Hypanis angusticostata, в олигогалинных водных массах Hypanis vitrea glabra.
Итак, формирование специфической каспийской фауны обусловлено наличием в прежние геологические эпохи специфической арены жизни – мезогалинных и олигогалинных водных масс и преодолением предками современной каспийской фауны мезогалинного барьера (=барьера критической солености). Преодоление этого барьера сопровождалось коренными перестройками не только осморегуляторных механизмов, но и различных морфологических признаков, что достаточно ярко обнаруживают эвригалинные морские виды в условиях современной регрессии Арала. Проникновение в новую адаптивную зону при наличии вакантных лицензий, что явно имело место при смене соленостного режима вследствие вымирания большей части фаун, могло обуславливать взрывной характер видообразования. Осолонение палеоводоемов влекло за собой сокращение объема и протяженности олигогалинных водных масс, сокращение этой специфической арены жизни и, следовательно, вымирание фауны или отступление (проникновение) отдельных форм в другую водную массу.
Показательно видообразование в олигогалинных и мезогалинных водных массах на примере таких специфических пресноводных видов, как легочные катушки рода Anisus. Современные Anisus обитают в пресных водах (Старобогатов, 1970), исключением являются четыре вида (Старобогатов, Ситникова, 1992), предки которых в отдаленные геологические эпохи, преодолев соленостные барьеры, проникли в олигогалинные, а в последующем и в мезогалинные водные массы. Распространяясь в соответствии с распределением пищевого материала первоначально, вероятно, в мелководных районах этих водных масс, Anisus радиируют, осваивая затем и глубоководные районы. Изменение солености, перемещение и, вероятно, уничтожение в результате неоднократно повторяющихся природных катаклизмов определенных водных масс вместе с населяющей их фауной привело к разрыву последовательного ряда видов, освоивших различные глубины в пределах олиго- и мезогалинных водных масс. Anisus в Каспии – это остатки древней запредельно специализированной группы. Иными причинами, нежели вымиранием промежуточных форм при уничтожении водных масс, сложно объяснить разрыв по градиенту солености, захватывающий два барьера, в распределении современных Anisus, обитающих и в пресных водных массах в бассейне современного Каспия, и на глубинах 30–200 м в пределах мезогалинных водных масс моря.
Водоемы Каспийской котловины (Апшеронское, Бакинское, Хазарское, Хвалынское и Новокаспийское, т.е. то, которое мы называем Каспийским морем) в той или иной мере преемственны по фауне (хотя комплексы родов и видов меняются) Акчагыльскому озеру-морю, в котором появилась богатая эндемичная фауна (Старобогатов, 1970, 1994).
С позиций концепции водных масс ясно, что вымирание специфической фауны той или иной водной массы могло проходить при небольшом повышении солености или кратковременном прекращении стока рек. Например, для Аральского моря, хотя оно и много меньше Каспия, потребовалось повышение средней солености воды менее, чем на 5 ‰ в течение всего 20 лет, чтобы оно полностью утратило специфическую фауну олигогалинных водных масс.
Учитывая значительные колебания уровня палеоводоемов, занимавших территорию современного Каспия и самого Каспия, нужно отметить, что вымирание специфической фауны той или иной водной массы, вероятно, происходило неоднократно, а при восстановлении водной массы было возможным быстрое проникновение видов из соседних водных масс, сопровождающееся бурным видообразованием.
В Каспийском море за время после хвалынского наиболее пострадала специфическая олигогалинная фауна, поскольку олигогалинные водные массы в нем в этот период занимали относительно небольшое пространство и резко меняли свои объемы даже в XX веке.
Создается впечатление (см. также Шорыгин, 1945, Мордухай-Болтовской, 1960), что современная мезогалинная фауна является выходцем из олигогалинных водных масс, лишь в недавнем прошлом преодолевшая хорогалинную зону. Как бы там ни было, специфическая мезогалинная фауна Каспийского моря приурочена к наименее минерализованной части мезогалинной водной массы и ее представители не отмечались в осолоненных заливах.
Фауна, существовавшая в гигантских озерах Паннонского, Эвксинского и Каспийского бассейнов, развивалась в условиях строгой изоляции от собственно (s. str.) морской фауны. В условиях «островной» изоляции возможны «запредельная специализация» и создание «монстров», которые при встрече с видами, развивавшимися на «магистральных путях эволюции» (Шварц, 1980) проигрывают в конкурентной борьбе. Так, после прорыва морских вод в Азово-Черноморском бассейне немногие уцелевшие представители понто-каспийской фауны оказались вытесненными в олигогалинные водные массы, причем им пришлось освоить и более пресные воды. Последнее обстоятельство позволило двум представителям двухстворчатых моллюсков с пелагической личиночной стадией (Dreissena bugensis и Hypanis colorata) в 20 веке расселиться в водохранилищах.
Проникновение или вселение морских эвригалинных видов приводит к сокращению ареалов или выпадению отдельных специфических представителей фауны в Каспийском море, последнем хранилище уникальной фауны древних озер-морей. Поэтому необходимо строго регламентировать акклиматизационные работы и максимально оградить Каспий от возможного завоза нежелательных вселенцев.
Предлагаемое деление всей гидросферы на шесть водных масс: пресноводные, олигогалинные, мезогалинные, эугалинные, гипергалинные и ультрагалинные основано на резком изменении физико-химических свойств воды вдоль градиента солености природных вод, к которым гидробионты по мере преодоления соленостных барьеров и освоения соответствующих водных масс должны вырабатывать соответствующие морфофизиологические адаптации. В пределах одной водной массы ареалы гидробионтов могут быть ограничены различными соленостными значениями. Так, в связи с длительным постоянством химического состава вод океана большое количество видов стеногалинны и не выносят даже незначительных колебаний океанических вод. Разумеется, стеногалинная фауна океана не ограничена никакими соленостными барьерами. Ее стеногалинность обусловлена, вероятно, лицензированием (см. Старобогатов, 1992) в процессе экспансии жизни в океане.
Континентальные водоемы различной минерализации лишены связи с океаном и проникновение пресноводных видов в олигогалинные, а затем и мезогалинные воды происходит при ослабленном ценотическом контроле со стороны немногих специфических видов, длительное время эволюционировавших в мезогалинных или олигогалинных водных массах.
Справедливо мнение В.П. Паули (1954) и Е. Даля (Dahl, 1956), считавших, что бедность фауны солоноватых водоемов обусловлена кратковременностью их существования в геологических временных масштабах. Если бы солоноватые водоемы в неизменном виде существовали периоды времени, соизмеримые со временем существования океана, то их жизнь по разнообразию была бы сравнима с жизнью океана. Так, согласно общей схеме распределения водной фауны в водоемах различной солености? максимальное число видов отмечается при средней солености океана и в пресных водах (рис. 8.10). В лишенных связи с океаном Каспийском и Аральском морях общее число видов меньше, чем в пресных водах, и, чем моложе водоем, тем меньше видов населяет его (рис. 8.11).
<![if !vml]>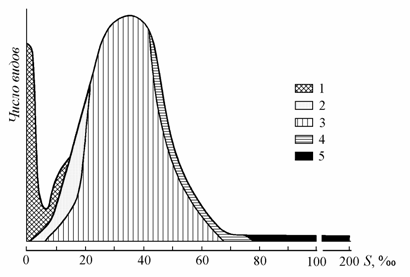 <![endif]>
<![endif]>
Рис. 8.10. Схема изменения облика водной фауны в водоемах различной солености (по: Хлебович, 1962; с дополнениями Kinne, 1971; из: Бергер, 1986). 1 – пресноводные, 2 – солоноватоводные, 3 – морские, 4 – гипергалинные, 5 – ультрагалинные виды.
<![if !vml]>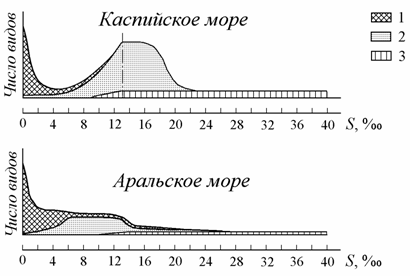 <![endif]>
<![endif]>
Рис. 8.11. Схема изменения облика водной фауны в Каспийском и Аральском морях (по: Андреева, Андреев, 2001б; левая часть схемы Каспийского моря заимствована из: Зенкевич, 1977). 1 – пресноводные, 2 – солоноватоводные, 3 – морские виды.
Вследствие наличия биотического барьера также затруднено проникновение и пресноводных видов в мезогалинные воды океанического состава при непосредственном впадении рек в океан или открытое море. В этом случае, как уже упоминалось выше, наименее заселенными оказываются олигогалинные водные массы и зона мезогалинного барьера. Первоначально на этот факт обратил внимание A. Ремане (Remane, 1934), а полный анализ распределения числа видов в водах океанического состава, в зависимости от фактора солености, привел В.В. Хлебович (1962, 1974). Признавая, вслед за В.В. Хлебовичем (1974), факт древнего вселения гидробионтов в пресные воды и дальнейшего их независимого развития, мы считаем (Андреева, Андреев, 2001б), что именно насыщенность пресных вод жизнью является в современных условиях мощным биотическим барьером на пути проникновения не только морских видов в пресные воды, но даже видов, длительно эволюционировавших в мезогалинных и олигогалинных водных массах. Для освоения пресных вод в современных условиях должно иметь место исключительное стечение обстоятельств, обеспечивающих проникновение видов. Для Dreissena bugensis и Hypanis colorata, помимо безусловного приобретения морфофизиологических адаптаций, обеспечивающих обитание в пресных водах, это постройка водохранилищ и завоз (возможно неоднократный) личинок или взрослых особей.
Итак, с одной стороны барьерные солености выступают как изолирующий фактор в распределении живого, препятствуя путем изоляции фаун в пределах определенной водной массы их смешению. Особенно велика роль барьерных соленостей в распределении осмоконформеров с пелагической стадией развития. Для организмов, обладающих более совершенными механизмами осморегуляции, они имеют, за исключением барьера критической солености, меньшее значение. Именно реальное существование барьерных соленостей явилось фактором, обеспечивающим формирование специфических и, часто, эндемичных фаун. С другой стороны, в пределах барьерных соленостей наблюдается резкое изменение физико-химических свойств вод, оказывающих, в частности, на двустворчатых моллюсков воздействие, вероятно, и как мощный мутагенный фактор.
Следует подробнее остановиться на механизмах, обуславливающих действие барьерных соленостей в качестве видообразующего фактора. Как уже отмечалось выше, в пределах соленостных барьеров (в частности для Аральского моря), помимо линейного нарастания солености, происходит скачкообразное изменение физико-химических свойств воды, важнейшим из которых, с точки зрения выживания морских эвригалинных двустворчатых моллюсков, является изменение соотношения ионов.
Установлено, что морские эвригалинные моллюски, являясь осмоконформерами, не способны активно регулировать осмотическое давление внутренней среды. В то же время их эвригалинность в пределах 5–8 — 75–80 ‰ обеспечивается морфофункциональными адаптациями на клеточном уровне. Обобщая литературные материалы и результаты собственных экспериментов, В.Я. Бергер (1976, 1979, 1986) указывает, что механизмы адаптаций морских эвригалинных моллюсков сложны и разнообразны, при этом универсальным механизмом клеточной адаптации к изменению солености среды обитания считается внутриклеточная изоосмическая регуляция, которая достигается как за счет низкомолекулярных азотсодержащих соединений (аминокислоты, таурин, бетаин, триметиламиноксид и др.), так и за счет изменения концентрации неорганических ионов, главным образом ионов натрия, и, вероятно, хлора. Ведущую роль в этих процессах имеет регуляция электролитного состава, обеспечивающая осмотическое выравнивание внутриклеточной и внешней среды.
Для морских эвригалинных моллюсков отмечены существенные перестройки электролитного состава внутри клетки при изменении солености окружающей среды. В частности, для морских моллюсков, впрочем, как и для других животных, характерно высокое содержание внутриклеточного калия, уровень которого снижается весьма незначительно как при акклимации к низкой солености, так и у моллюсков, существующих в солоноватых водоемах. В то время как содержание натрия в клетке варьирует в весьма значительных пределах. Широкое варьирование содержания иона натрия в клетке позволяет рассматривать его в качестве главного агента различных сторон жизнедеятельности клетки при изменении среды, в частности, установлено, что изменения концентрации ионов натрия могут эффективно влиять на реализацию генетической информации (Хлебович, 1974; Бергер, 1986; и др.). Возможно, с этим связаны, отмечаемые В.Я. Бергером и А.Д. Харазовой (1971, 1977), особенности изменения синтеза РНК и белка в клетках различных тканей моллюсков при смене солености среды. Предполагается (Бергер, 1986), что угнетение биосинтетической деятельности является следствием нарушения конформации, необходимой для нормального функционирования молекул РНК и белков. Можно полагать, что скачкообразное изменение соотношения ионов в пределах барьерных соленостей обуславливает еще более существенное нарушение процессов реализации генетической информации. Доказательством этого положения может служить колоссально возросшая изменчивость эвригалинных двустворчатых моллюсков при преодолении экосистемой Аральского моря барьерных соленостей (Андреева, 2000б).
Итак, изменение солености среды обитания эвригалинных морских моллюсков может эффективно влиять на реализацию генетической информации. На основе наблюдений за морфологической изменчивостью двустворок Аральского моря при повышении солености его вод можно с уверенностью утверждать, что именно резкое изменение физико-химических свойств воды (в том числе и ионного состава) в пределах барьерных соленостей явилось причиной нарушения процессов воспроизводства генетической информации, приведшего к превышению не только внутривидовых, но и межвидовых пределов изменчивости жизненно важных морфологических признаков. Вероятно, эти процессы универсальны для всех водоемов с соответствующей соленостью воды. Так, повышенную ненаправленную изменчивость при гидрологических кризисах (Парамонова, 1971; Невесская с соавт., 1986) обнаруживают сарматские кардииды и другие моллюски морских и солоноватых палеоводоемов.
Таким образом, в силу нарушения воспроизводства генетической информации в пределах соленостных барьеров, последние выступают в качестве факторов, вызывающих взрыв ненаправленной изменчивости двустворчатых моллюсков, благодаря чему появляется массовый материал для эволюционного процесса. С другой стороны, для осмоконформеров все соленостные барьеры являются изолирующим фактором, обеспечивающим их независимую эволюцию в пределах водных масс, отсекаемых соленостными барьерами. Для осморегуляторов наиболее существенным является мезогалинный барьер. Эволюционные процессы, сопровождающие последний гидрологический кризис современного Аральского моря, являются моделью эволюционных преобразований фаун водоемов, существовавших в геологическом прошлом.