
Проблемы Эволюции |
М.: Наука, 1980
<![if !supportEmptyParas]> <![endif]>
{149}
Глава V
ЭКОЛОГИЧЕСКИЕ МЕХАНИЗМЫ ПОДДЕРЖАНИЯ ГЕНЕТИЧЕСКОЙ РАЗНОРОДНОСТИ ПОПУЛЯЦИЙ
Когда генетическая разнородность популяции принимает характер выраженного полиморфизма, ее биологическое значение становится очевидным. Оно заключается в большом диапазоне условий, которые популяция в целом может использовать для поддержания оптимальной численности. Приведенные в главах II и IV данные и их обсуждение показывают, что современная экология еще далека от понимания роли полиморфизма в жизни животных. Степень приспособленности разных генотипов к разным условиям среды достигает такой степени специфичности, о которой мы сейчас можем только догадываться. Об этом говорят некоторые экспериментальные данные, свидетельствующие о том, что не только представители разных фаз, но и разные генотипы занимают различное положение в системе «популяция — среда», так как занимают различные экологические субниши [Lewontin, 1955]. В пользу развиваемых Левонтиным представлений свидетельствуют убедительные косвенные данные. Оказалось, что популяции с выраженным хромосомным полиморфизмом отличаются более широким распространением и большим разнообразием занимаемых биотопов [Cunha, Dobzhansky, 1954; Cunha et al., 1959], а для популяции с обедненным генофондом характерно ограниченное число занимаемых экологических ниш.
По-видимому, полную ясность в этот важнейший вопрос эволюционной теории может внести лишь углубленное экологическое исследование, позволяющее связать особенности отдельных популяций не только с их общими генетическими особенностями, но и степенью генетической разнородности. Пока такие исследования, по существу, отсутствуют, необходимо ограничиться самым общим утверждением: повышение генетической разнородности популяции выгодно, так как способствует более полному использованию ресурсов среды. С другой стороны, обогащение генофонда популяции резко повышает возможности ее приспособительного преобразования как единого целого, резко повышает ее жизнеспособность и по существу является гарантией ее существования в меняющихся условиях среды. Как будет показано ниже, тот же самый механизм, который обеспечивает приспособление популяции к изменениям условий среды ее родины, создает предпосылки к расширению ареала. Отсюда понятно, что в природе должны существовать
{150}
разнообразные механизмы поддержания генетической разнородности популяции и непрерывного обогащения ее генофонда.
Общее обогащение генофонда популяции не только повышает приспособительные возможности популяции в целом, но и увеличивает генетическую емкость отдельных особей (гл. II). В этом отношении особый интерес представляют эксперименты Кэрсон [Carson, 1961]. Оказалось, что популяция, происходящая от одной пары Drosophila robusta из центра ее ареала, по степени своей жизнеспособности и способности к дальнейшим приспособительным преобразованиям не отличается от популяций, возникших от большого числа исходных производителей (основателей). Опыты Кэрсон подкупают тщательностью выполнения, но, по существу, они лишь подтверждают многие данные, полученные ранее. Известно, что лабораторные колонии ряда диких видов животных происходят от немногих основателей. В нашей лаборатории основателями крупных колоний полевки Миддендорфа, экономки, пеструшки, узкочерепной полевки были всего несколько пар особей; несколько сот полевок Миддендорфа получены от одной пары.
Популяции многих акклиматизированных видов происходят от ничтожного числа особей-основателей. К. Драчевский [1961] указывает, что в Киргизии от трех пар нутрий за 4 года было получено процветающее стадо в 300 голов. В Англии популяция нутрии возникла за счет отдельных зверьков, сбежавших со звероферм. Вряд ли таких особей могло быть много. Тем не менее за 10 лет нутрия распространилась на территории нескольких графств, а в 1961–1962 гг. было добыто 100 тыс. животных [Norris, 1963]. Три пары лосей, выпущенных на о-ве Ньюфаундленд, дали процветающую популяцию, численностью 30–40 тыс. голов [Pimlott, 1961]. Канадская популяция зайца-русака берет начало от семи самок и двух самцов, вывезенных из Германии в 1912 г. [Dean, De Vos, 1965]. От 14 кабанов, отловленных в 1909 г. в России, возникла популяция кабанов в США. Территория, которую она сейчас занимает, превышает 1000 км2 [Иванов, 1962]. Подобных примеров известно много. В обобщающей статье А. А. Насимович [1961] пишет: «Многочисленны примеры, когда выпуск небольшого количества животных приводил к быстрому размножению и последующему распространению на огромной территории интродуцированного вида (кролики, ондатра, домовой воробей).
Нами [Шварц и др., 1966] был поставлен специальный эксперимент, цель которого исследовать, как меняется изменчивость популяции при ее формировании на основе немногих случайных основателей. Мы сравнивали изменчивость окраски грызунов из природных популяций и из лабораторных колоний. Окраска определялась колориметрически, что гарантирует объективность ее оценки. Основателями колоний были животные тех же популяций, которые служили нам контролем: полевки Миддендорфа (Microtus middendorffi) — 2 самца и одна самка, экономка (М. оесоnо-mus) — 6 самцов и 4 самки, северная узкочерепная полевка (М. g.
{151}
major) — 5 пар, М. g. gregalis — 4 самца и 6 самок. Животные разводились в течение 4–6 поколений. Было установлено, что ни в одном из 4 случаев существенного изменения диапазона и направления варьирования не произошло, принцип основателя не сработал. В свете результатов опытов Кэрсон [Carson, 1961] это не кажется нам удивительным. Мы имели возможность показать, что популяция, возникшая от немногих случайных основателей, полностью сохраняет способность к направленным изменениям под влиянием отбора. Об этом свидетельствует специальный эксперимент [Шварц, Покровский, 1966], результаты которого обсуждаются нами в этой книге.
О генетической емкости отдельных индивидов и, следовательно, о возможной генетической разнородности популяций, возникших от немногих основателей, свидетельствуют также иммунологические исследования трансплантационных антигенов. В нашей лаборатории Л. М. Сюзюмова изучала сроки отторжения трансплантатов кожи в разных группах полевки-экономки. В тех случаях, когда реципиент и донор относятся к разным подвидам (М. oeconomus oeconomus, М. о. chahlovi), сроки отторжения трансплантата варьируют от 6 до 7 дней. При пересадке кожного лоскута в пределах одной семьи (реципиент и донор — потомство одной самки) средний срок жизни трансплантата, естественно, увеличивается (колебания от 6 до 30 дней). Уже эти опыты показывают, что в пределах близкородственных групп животных (одна семья) можно обнаружить антигенные, а следовательно, и генетические различия, соизмеримые с различиями между подвидами. Л. М. Сюзюмова искусственно увеличила генетическую разнородность экспериментальной колонии путем гибридизации изучаемых подвидов. Обогащение генофонда подопытной группы животных привело к тому, что при внутрисемейной трансплантации (от брата к брату) продолжительность жизни пересаженных лоскутов кожи нередко еще более сокращается. Так, в пределах одной из гибридных смесей срок жизни трансплантатов колебался от 6 до 8 дней. В других семьях наблюдалась большая изменчивость в проявлении реакции на трансплантат (срок жизни пересаженного лоскута от 7 до 52 дней). Кажется очевидным, что популяция, возникающая на основе размножения подобных семей, может быть генетически столь же разнородной, что и процветающие природные популяции. В этих условиях принцип основателя может иметь лишь ограниченное значение.
Все эти, а также громадное количество других фактов показывает, что обогащение общего генофонда популяции имеет огромное биологическое значение. Естественно поэтому ожидать специальных механизмов поддержания в популяции генетического разнообразия. Один из таких механизмов — повышенная жизнеспособность гетерозигот.
Этот вопрос подробно разбирается в генетической литературе, и здесь нет нужды его детально анализировать. Важен сам факт:
{152}
гетерозиготы в большинстве случаев обладают большей жизнеспособностью, что содействует сохранению в популяции генов, которые в гомозиготном состоянии являются вредными или даже летальными. Конкретные причины повышенной жизнеспособности гетерозигот не могут считаться выясненными. Существуют данные, показывающие, что важна не гетерозиготность сама па себе, а вполне определенное сочетание генов. Об этом свидетельствуют, в частности, эксперименты Уоллеса [Wallace, 1955], показавшего, что среди гетерозигот единой популяции обнаруживаются все степени жизнеспособности вплоть до особей, отличающихся малой жизненностью. С другой стороны, имеются наблюдения, указывающие на специфические частные причины повышенной выживаемости гетерозигот. Лучший пример — распространение среди людей серповидного гемоглобина (Нbs). Гомозиготы НbsНbs, как правило, не доживают до половой зрелости. Тем не менее в ряде местностей Африки частота встречаемости гена Нbs достигает 20%. Аллисон [Allison, 1956] установил, что это связано с повышенной сопротивляемостью гетерозиготных индивидуумов к малярии. Оказалось далее, что Нbs встречается и в других частях земного шара, где малярия является существенным фактором смертности. Эти наблюдения приводят к выводу, что отклонения от нормального типа гемоглобина ухудшают условия развития паразита и снижают таким путем общий процент смертности среди местного населения, несмотря на значительную смертность гомозигот HbsHbs. Весьма вероятно, что сходным образом объясняется и распространение в популяциях гетерозигот иных типов. Ясно, однако, что чем выше генетическая разнородность популяции, тем больше возможности для возникновения наиболее благоприятных вариантов. Это оправдывает изыскание механизмов поддержания в популяции высокой генетической изменчивости.
Несомненно, что одним из ведущих механизмов поддержания гетерозиготности популяции и общего обогащения генофонда являются все те же механизмы, которые способствуют перемешиванию популяций и микропопуляций. Примеры этого порядка очень хорошо известны и поэтому мы ограничимся лишь немногими.
У большинства видов млекопитающих хорошо выражена сезонная смена мест обитания. Так, водяная полевка осенью покидает сплавинные острова и частично берега озер и на зиму поселяется на лугах и в стогах сена. У ондатры в период гона наблюдаются массовые перекочевки, оканчивающиеся обычно со спадом воды. Кочевки часто имеют следствием заселение ондатрой новых водоемов и несомненно приводят к постоянному перемешиванию популяций. Многие мелкие виды мышевидных грызунов в степных и лесостепных областях весной и в первую половину лета занимают самые разнообразные биотопы: колки, заросли кустарников, посевы культурных растений, залежи и т. д. С наступлением летней жары большинство из них покидают эти
{153}
места обитания и поселяются по берегам водоемов. Это безусловно приводит к скрещиванию особей из различных мест обитания.
В лесостепи Зауралья нами установлено наличие резко выраженных миграций у обыкновенной бурозубки. Летом в условиях засушливого климата землеройки этого вида встречаются только по берегам водоемов. С наступлением осенней погоды связь их с увлажненными местообитаниями делается менее тесной, и они занимают самые разнообразные биотопы в степи и лесостепи. При концентрации их у водоемов следующим летом смешение популяций неизбежно. Лоси оставляют зимой излюбленные летом болота и часто придерживаются ограниченных площадей приречных древостоев. Песец и северный олень осенью покидают летние места обитания в тундре и зимуют в лесотундре или даже в северных частях лесной зоны. Сотрудник нашей лаборатории В. С. Смирнов проводил кольцевание песцов на Ямале. Один из зверей через несколько месяцев был добыт на Аляске. Дзерен зимой появляется в степях Забайкалья и Северной Монголии, а весной возвращается обратно в Южную Монголию. Ежегодные миграции на громадные расстояния совершают котики, моржи, киты, летучие мыши и другие млекопитающие.
Миграции, а тем более миграции на далекие расстояния неизбежно увеличивают вероятность спаривания животных из разных популяций. Как известно, для птиц миграции еще более характерны, чем для млекопитающих. Напомним также, что миграции известны и у рептилий; так, они наблюдались у обыкновенных ужей [Терентьев, Чернов, 1949], узорчатого полоза [Хозацкий, Эглон, 1947], прыткой ящерицы [Щепотьев, 1952].
Говоря о механизме повышения гетерозиготности, необходимо отметить особое значение расселения молодых при переходе их к самостоятельному образу жизни. Это явление хорошо изучено для относительно немногих видов, но, вероятно, имеет широкое распространение.
Н. И. Калабуховым и В. В. Раевским [1935] установлено, что у малого суслика к передвижению более всего склонны молодые животные. В. Н. Павлинин [1948] при помощи кольцевания установил, что молодые кроты имеют тенденцию отходить на большое расстояние от места своего рождения. Из 112 вновь пойманных окольцованных кротов на расстоянии 50–500 м от точки выпуска было добыто: взрослых — 79%, молодых — 58%. На расстоянии свыше 500 м были встречены только молодые особи. Среди пойманных на участке кольцевания кротов взрослых меченых оказалось 71; а молодых только 47%.
А. А. Насимович и соавторы [1948], разбирая вопрос о миграциях норвежского лемминга, пишут: «Особенно в большом числе из поселений эмигрируют молодые и подростки; поэтому летне-осенние миграции леммингов прежде всего являются расселением молодых». О расселении молодых соболей из района материнского гнезда свидетельствуют материалы В. В. Раевского [1947]. У не-
{154}
которых млекопитающих половое созревание наступает в строго определенном возрасте и относительно очень поздно (бобр, сурок, обыкновенный хомяк, крот, землеройки). Землеройки, например, живут 13–15 месяцев, а половая зрелость наступает у них только в возрасте 8–10 месяцев. Сравнительно длительный период полового созревания благоприятствует более широкому расселению молодых особей со всеми вытекающими отсюда биологическими последствиями.
Особо следует отметить, что у ряда видов особи разного пола и возраста имеют или разные районы зимовок, или покидают их в разные сроки. У таких видов самки и самцы, составляющие единую популяцию, в какие-то периоды подвергаются воздействию различных внешних условий. У северного оленя старые самцы и яловые самки уходят зимой на юг дальше, чем другие особи. У котика первыми приходят весной к Командорским островам секачи, а через 3–4 недели неполовозрелые самцы, позднее всех прибывают неполовозрелые самки. У кашалотов самки почти не выходят за пределы тропиков и субтропиков, а самцы в теплые месяцы года откочевывают на север до Баренцева моря и берегов Камчатки [Томилин, 1938]. В Таджикистане кабаны зиму и весну проводят в пойменных зарослях. Молодые животные и средневозрастные самцы на лето откочевывают в горы, а матки с поросятами и старые одинцы остаются в пойме. У ряда видов горных баранов самки с молодняком держатся на сравнительно небольших высотах, взрослые же самцы и подросший молодняк обитают в верхних зонах, доходя до высокогорий [Цалкин, 1945].
Аналогичные примеры мог бы привести любой зоолог из своей практики. Из них следует очень важный вывод, что, ведя различный образ жизни, самцы и самки неминуемо подвергаются различным силам отбора. В силу этого их генетический состав не может быть одинаковым, что с неизбежностью ведет к общему увеличению генетической разнородности популяции. С этой точки зрения новое освещение получают и хорошо известные факты резких экологических и морфофизиологических различий между самцами и самками, которые особенно хорошо выражены у рыб.
Известно, что у очень многих видов рыб (лососевые, глубоководные удильщики, некоторые губаны и др.) в популяции наряду с нормальными самцами имеются и карликовые самцы, резко отличающиеся и по морфологии, и по своему образу жизни. Лучший пример — жилые карликовые самцы у проходных видов сиговых. Нормальная воспроизводительная функция карликовых самцов доказана, их численность колеблется в зависимости от условий среды [Европейцева, 1962] и коррелирована с численностью проходных самцов. Средняя относительная численность самцов, видимо, фиксирована генетическими механизмами, так как они составляют довольно значительный процент молоди, выращенной в искусственных условиях (наблюдения на куринском лососе). Естественно, что карликовые самцы и нормальные самки подвер-
{155}
жены совершенно разным силам отбора, их генетический состав не может не быть различным. В силу этого присутствие в популяции карликовых самцов поддерживает генетическую разнородность популяции, увеличивает богатство того котла, из которого отбор черпает ресурсы для непрерывного совершенствования живых организмов 1.
Для подвижных животных миграции и перемещения, несомненно, являются одним из основных механизмов поддержания генетической разнородности популяций и тормозом обеднения общего генофонда. Однако известно большое число малоподвижных животных, для которых этот механизм может иметь лишь ограниченное значение. Здесь вступает в силу другой механизм, сущность которого сводится к тому, что пары формируются за счет животных, принадлежащих к разным поколениям. Так как (подробно см. гл. VI) генетическая структура разных поколений различна, то конечный эффект этого механизма должен быть весьма сходным с миграционным механизмом поддержания генетической разнородности.
Как известно, у целого ряда млекопитающих в период гона наблюдаются бои самцов за самку. При этом половозрелые молодые самцы не принимают участия в размножении до тех пор, пока не достигнут полного физиологического расцвета и смогут противостоять самцам зрелого возраста. Несколько примеров поясняют сказанное.
Самки зубра достигают половой зрелости на 3–4-й год жизни, самцы — на 4-й, но вступают в размножение не раньше чем в возрасте 6–7 лет, так как отгоняются более старыми самцами. Самцы и самки оленей созревают примерно в одном возрасте, но самцы принимают участие в размножении много позднее. Сходное наблюдается и у кабанов: самцы и самки созревают примерно одновременно, но молодые самцы допускаются к размножению, только достигнув 6–7 лет. Относительно сибирского горного козла В. И. Цалкин [1950] пишет следующее: «Половозрелости самки достигают в возрасте около полутора лет и в двухлетнем возрасте уже часто имеют молодых. Половозрелость самцов наступает примерно в то же время. Однако непосредственное участие в размножении они начинают принимать лишь значительно позднее, так как отгоняются более крупными и старыми самцами». Вполне аналогичное явление наблюдается и у ластоногих. Самки морского котика размножаются с 3-летнего возраста, в то же время насту-
1 Наличие в популяции карликовых самцов может иметь и иное биологическое значение — экономию кормовых ресурсов популяции. Однако, если этот фактор и имеет значение, роль карликовых самцов в поддержании разнородности популяции остается несомненной. Какой фактор первичен, какой вторичен — не столь уж важно. Могут иметь значение и оба (экономия корма и поддержание генетической разнородности). В таком случае мы сталкиваемся со своеобразной мультифункциональностью экологической особенности вида.
{156}
пает и половое созревание самцов. Однако до 7 лет самцы котика в размножение не вступают и занимают отдельные лежбища. Борьба самцов за самку, хотя и в менее выраженной форме, имеет место и у ряда хищных зверей (волк, медведь, тигр), что дает основание полагать, что и у них производителями являются преимущественно самцы, уже достигшие расцвета своих сил.
Все сказанное свидетельствует о том, что у большой группы млекопитающих самцы младшего возраста не принимают участия в размножении, а молодые самки кроются самцами старшего возраста. С другой стороны, многочисленными наблюдениями показано, что старые самцы, сохранившие еще половую потенцию, бывают вынуждены уступить свое место более молодым производителям. Это приводит к тому, что наиболее старые самцы исключаются из размножения, подобно наиболее молодым. В силу большей естественной смертности взрослых самцов по сравнению со взрослыми самками средняя продолжительность их жизни, как правило, меньше, чем у самок.
Сходное явление было недавно описано на тетеревах А. М. Чельцовым-Бебутовым [1965] под названием «возрастного кросса». Автор показал, что сложный характер тетеревиных токов приводит не к беспорядочному спариванию, а «к преимущественным встречам определенных и, что особенно важно, разнородных в физиологическом отношении групп популяции: менее активные, изгнанные с тока самцы имеют больше шансов оплодотворить наиболее активных в половом отношении самок, тогда как менее активные, имеющие меньший порог раздражения самки чаще спариваются с наиболее сильными и активными косачами, остающимися «победителями на току». Сопоставляя «активность» птиц с их возрастом, автор приходит к очень важному заключению, что тока тетеревиных обеспечивают преимущественное спаривание взрослых самок с молодыми самцами, а молодых тетерок — с петухами старших возрастов. А. М. Чельцов-Бебутов полагает, что возрастной кросс имеет в своей основе повышение жизненности потомства, полученного от родителей разного возраста. Это предположение не лишено оснований, так как подтверждается некоторыми наблюдениями над домашними животными [Вахрушев, Волков, 1945; Милованов, 1950; Старков, 1952; Бригес, 1953; Анорова, 1959, 1960, 1964; и др.]. Однако каковы бы ни были первопричины возрастного кросса, его важнейшее генетическое следствие сводится к увеличению генетической разнородности популяций.
Более сложен вопрос о закономерностях, которым подчиняется подбор пар у животных, не обладающих половым диморфизмом и для которых борьба самцов за самку не характерна. Казалось бы, что в этом случае таких закономерностей вообще нет и все дело решает случай. Наши наблюдения над грызунами показали, что это не так. Известно, что самки грызунов созревают несколько раньше самцов. К чему это приводит, покажем на конкретном примере.
{157}
В условиях лесостепного Зауралья, где проводились наши исследования, первый помет водяных полевок рождается в начале мая или (в отдельные годы) в конце апреля. Самки этой генерация созревают в первой половине июня. В это время молодые самцы еще не половозрелы, и поэтому молодые самки могут быть покрыты только самцами старшего возраста (перезимовавшими), что и в действительности имеет место, так как в то время, когда молодые самцы еще остаются неполовозрелыми, подавляющее большинство самок оказываются беременными. Так, например, в 1951 г. в Курганской области первый половозрелый самец текущего года рождения был добыт 15 июня. Между тем беременные самки добывались уже в первой декаде июня, а в последних числах мая было добыто несколько молодых самок в состоянии течки. Ко времени созревания молодых самцов часть молодых самок уже выкармливали молодняк. Так, 15 июня была добыта молодая самка, у которой беременность сочеталась с лактацией. Аналогичное явление наблюдалось в Курганской области и в 1950 г. Половозрелые молодые самцы начали попадаться только с 19-го июня, между тем как уже в конце первой декады июня часть молодых самок была беременна вторично (сочетание лактации и беременности). Нет сомнения, что созревшие самцы немедленно вступают в размножение, так как перезимовавших (старых) самцов в это время уже настолько мало, что если бы молодые самцы не выступали уже в роли производителей, это привело бы к массовому прохолостанию самок, чего в действительности не происходит. Однако первое время после созревания молодых самцов самки этой генерации уже беременны. Следовательно, молодые самцы кроют преимущественно перезимовавших самок, которые в это время уже с полным основанием могут быть названы старыми. Водяные полевки второго помета рождаются в июне. Далеко не все особи этой генерации успевают достичь половой зрелости в год рождения, преимущественно самки. Следовательно, и в этом случае молодые-самки кроются самцами старшего возраста.
Наблюдения показали, что совершенно такая же картина выявляется и при анализе «состава пар» и у других видов грызунов (полевка-экономка, узкочерепная полевка, красная полевка и др.). Согласно данным А. А. Слудского [1948], основанным на изучении очень большого материала, самцы ондатры в год своего рождения никогда не вызревают, в то время как часть самок приносит помет в возрасте не более 4 месяцев. Естественно, что они могут быть покрыты только более старыми самцами. Наши несравненно более скромные данные подтверждают выводы А. А. Слудского для северной лесостепи Зауралья.
Приведенные примеры показывают, что разная скорость полового созревания приводит к тому, что пары формируются преимущественно за счет животных разного возраста, разных генераций. Следствием этого является непрерывное восстановление генофонда популяции, даже в том случае, если в отдельных генерациях гено-
{158}
фонд окажется существенно нарушенным. Кажется в высшей степени показательным, что у всех животных скорость полового созревания самцов и самок различна. Приведенные нами примеры касаются млекопитающих. Разная скорость полового созревания рыб отмечается Г. В. Никольским [1965] как хорошо известный факт. Он приводит и ряд конкретных примеров, ясно показывающих, что формирование пар из животных разных генераций в ряде случаев становится неизбежным. «Так, например, у обыкновенного карася в исключительно благоприятных условиях роста разница во времени созревания самцов и самок сглаживается (в обычных условиях самцы созревают несколько раньше самок), и оба пола созревают в одном возрасте 1+ [Schaperclaus, 1953]. При этом относительная численность самцов уменьшается среди младших возрастных групп, но возрастает среди старших». В этом наблюдении обращает на себя внимание, что условия для формирования одновозрастных пар создаются лишь в наиболее благоприятной среде. Возможно, это правило распространяется и на других животных. Его биологический смысл с развиваемых здесь позиций ясен. Поддержанию генетической разновидности популяций у рыб, безусловно, способствует и разновозрастность полового созревания животных одного поколения.
Вряд ли можно сомневаться в том, что столь широкое распространение такой важной биологической закономерности, как различная скорость созревания полов, случайно. Законно полагать, что в основе этого явления, которое почти столь же всеобще, как сексуализация почти всех живых существ, лежат фундаментальные биологические закономерности. Приведенный анализ фактов делает вероятным, что это фундаментальная биологическая закономерность — механизм формирования пар, исключающий возможность обеднения генофонда популяции животных.
Возможно, что поддержание генетической разнородности в некоторых случаях определяется и этологическими механизмами. В этом вопросе еще очень много неясного, но постепенно накапливаются материалы, показывающие, что выбор брачных партнеров не подчиняется простой случайности. Это распространяется не только на животных, для которых характерны турнирные бои за самку. Избирательное спаривание обнаружено у простейших [Jennings, 1911], насекомых [Tower, 1906; Petit, 1956], амфибий [Sawada, 1963], птиц, [O'Donald, 1959], млекопитающих [Frederickson, Birnbaum, 1956; Mainardi, 1964; Levine, Lasher, 1965; и др.]. Большинство наблюдений свидетельствует о том, что избирательное спаривание осуществляется на основе морфологического сходства. Это проявляется, в частности, и в том, что при возможности выбора пары формируются за счет животных одного, а не разных подвидов [Mainardi et al., 1965]. Однако более детальное исследование того же Майнарди [Mainardi, 1964; Mainardi et al., 1965] показало, что в пределах подвида самки предпочитают спариваться с генетически неродственными самцами. Сделана первая попытка
{159}
оценить значение избирательного спаривания для развития популяции с помощью счетных машин. Были получены данные, показывающие, что, если брачные партнеры сходны, фиксируется один из аллелей (обычно доминантный), в противоположном случае устанавливается стабильный полиморфизм [Mainardi et al., 1966]. Не менее интересны наблюдения, показывающие, что сочетание разных генотипов в формирующихся парах у дрозофилы изменяется при изменении внешней среды [Petit, 1956]. Возможно, что расширение исследований в этом направлении позволит выявить новые механизмы поддержания в популяции генетической разнородности. Самцы и самки отличны не только физиологически, но и экологически, различно их отношение к среде, которое особенно отчетливо проявляется в различной смертности. Отметим лишь несколько наиболее интересных наблюдений в этой области. На основании изучения литературного материала по соотношению полов у 54 видов и 12 пород млекопитающих Б. С. Кубанцев [1964] пришел к выводу, что у большинства видов самцов рождается больше, чем самок. Известно, однако, что в большинстве популяций среди взрослых животных соотношение полов близко 1:1. Это само по себе свидетельствует о дифференцированной смертности животных разного пола. Повышенная смертность самцов по-разному проявляется в разных условиях среды. На чернохвостом олене (Odicoileus hemionus columbianus) показано, что уже в самом младшем возрасте самцы гибнут значительно чаще самок [Taber, Dasmann, 1954]. Это объясняется их большей активностью, которая, в свою очередь, определяется более высоким уровнем обмена. Поэтому при ухудшении условий соотношение полов резко сдвигается в сторону самок. Неблагоприятное воздействие низкой температуры вызывает повышение смертности половозрелых самцов мышей, но среди молодых различий в выживаемости не наблюдается [Zarron, Denison, 1956]. Относительно более высокая смертность наблюдается также при резком вмешательстве в жизнь популяции, которое происходит, например, при борьбе с вредителями сельского хозяйства. Опыты Н. И. Калабухова [1944] показали, что при химической обработке земель, зараженных обыкновенной полевкой (Microtus arvalis), остающиеся в живых грызуны (30–50%) —в основном самки, большинство из которых беременные и кормящие. Позднее Н. И. Калабухов с сотрудниками [1950] показали, что беременные самки малого суслика (Citellus pygmaeus) неохотно берут отравленную приманку, и смертность их при химической обработке зараженных территорий оказывается неизмеримо меньшей, чем смертность самцов и активного молодняка.
У некоторых видов закономерное превышение смертности самцов над смертностью самок приводит к возникновению определенных механизмов, обеспечивающих численное доминирование самцов при рождении. Это наблюдается, например, у ондатры. Согласно наблюдениям А. К. Околовича и Г. К. Корсакова [1951],
{160}
во многих районах соотношение полов при рождении приближается к 60:40.
Не следует, однако, думать, что смертность самцов всегда выше, чем смертность самок. Резко повышенная смертность самок наблюдается, например, на зимовках некоторых видов летучих мышей (Pipistrellus subflavus), что приводит к снижению их относительного количества до 20% [Davis, 1959]. Еще более интересно, что самки иногда оказываются более чувствительными к неблагоприятным условиям уже па эмбриональной стадии развития, что, естественно, приводит к численному преобладанию самцов среди молодняка [Zimmermann, 1963, наблюдения на шиншилле].
Изменения условий среды могут, по-видимому, вызывать изменение соотношения полов в популяции не только путем изменения относительной смертности животных разного пола, но и путем изменения соотношения полов при рождении. Влияние среды на соотношение полов может быть как косвенным, так и прямым. Специальный симпозиум, посвященный проблеме «возраст родителей и потомство», показал, что возраст родителей в значительной степени определяет свойства потомства [Cowary, 1954; итоги симпозиума см. также Uda, 1957]. Естественно, что косвенно это влияет и на соотношение полов. Становится ясным, что возрастная структура популяции может оказать влияние на соотношение полов среди молодых животных. Прямое влияние среды на соотношение полов имеет в своей основе разную реакцию половых хромосом на изменение биохимизма организма [Uda, 1957]. В частности, доказано, что рН крови оказывает на соотношение полов настолько сильное влияние, что создает предпосылки для регулирования в потомстве числа самцов путем селекции [Whirter, 1956]. О различной реакции Х- и Y-хромосом на внешние воздействия свидетельствуют и известные эксперименты Б. Л. Астаурова [обзор см. Астауров, 1963] по регуляции пола у шелкопряда.
Указанные наблюдения делают понятным влияние на соотношение полов такого фактора внешней среды, как изменение режима питания, которое обнаруживается даже у людей [Uda, 1957], а у домашних животных наблюдалось неоднократно [Милованов, 1950; Лысов, Письменная, 1951; Аверьянов и др., 1952; Лукина, 1953; и др.].
Об исключительно большом значении регуляции определенного соотношения самцов и самок свидетельствует возникновение сложнейших генетических механизмов регуляции полового соотношения, которое наблюдается у некоторых животных. Исследование с ракообразными (Copepoda) показали [Battaglia, 1965], что у Tisbe reticulata снижение гетерогенности популяции неизбежно ведет к снижению относительного числа самок. Экологически это, естественно, объясняется тем, что при высокой плотности вероятность оплодотворения самок велика даже при низкой численности
{161}
самцов (Т. reticulata — полигамы). При снижении плотности популяции возникает опасность прохолостания части самок, их количество увеличивается. Эксперименты показали, что при близкородственном разведении Т. reticulata отбор на увеличение относительного числа самцов оказывается эффективным, увеличить же число самок в этих условиях оказывается невозможным. Батаглиа удалось установить не только экологический, но и генетический смысл этого явления. Оказалось, что у копепод пол определяется полифакторально: самки детерминируются несколькими доминантными генами, самцы — рецессивными. Поэтому число самцов определяется степенью гомозиготности популяции. При снижении численности происходит снижение генетической разнородности популяции, которая сопровождается относительным увеличением численности самцов, что, по изложенным выше причинам, снижает возможность прохолостания самок. Таким образом, у Copepoda выработался четкий генетический механизм, определяющий оптимальное соотношение между плотностью популяции, ее генетической структурой и соотношением полов. Весьма вероятно, что аналогичные механизмы существуют и у других животных, в том числе и у позвоночных. Об этом, в частности, свидетельствует интересная работа Стера [Stehr, 1964], специально изучавшего роль конкретных механизмов определения пола в микроэволюции. К этим исследованиям примыкают наблюдения, показывающие, что для генетически различных лабораторных популяций характерно различное соотношение полов [Levy, 1965].
Естественно, что дифференцированная смертность и различия в соотношении полов при рождении должны иметь следствием и различия в динамике численности животных разного пола, и различия в динамике структуры их генетического состава. Это делают очевидным экспериментальные исследования К. Петрусевича [Petrusewicz, 1958], показавшего, что при равном соотношении полов при рождении и повышенной смертности самцов в лабораторных популяциях мышей скорость изменений численности п амплитуда ее колебаний у самцов оказались выше в 43 случаях из 47.
Различная динамика численности должна иметь следствием различный генетический состав самцов и самок. Отсюда следует, что поддержание оптимального соотношения полов является вместе с тем и важнейшим механизмом поддержания генетической разнородности популяции. Нет достаточных оснований говорить о специальной роли самцов и самок в этом процессе, как это делает В. А. Геодакян [1965], приписывающий самцам ответственность за качество, самкам — за количество потомства, но можно с полным правом утверждать, что самцы и самки — это две не только физиологически, но и генетически различные группы животных. Поэтому можно согласиться с авторами, проводящими аналогию между разными клонами одноклеточных организмов, размножающихся половым путем, и полами высших животных [Kallmus,
{162}
Smith, 1960]. Отсюда вытекает необходимость экологического анализа последствий нарушения нормального соотношения полов. Исследование этого вопроса находится еще в самом начале, но уже имеющиеся данные ясно показывают, что экологические механизмы играют ведущую роль в поддержании генетической разнородности популяции. С другой стороны, значение этих механизмов делает понятным ряд общебиологических явлений широчайшего распространения (сексуализация всего живого мира, разная скорость созревания самцов и самок, в том числе и в их крайнем проявлении — карликовость самцов и т. д.).
Эти заключения приводят к выводу и более общего характера, касающегося значения генетической разнородности популяций. Разделение вида на два пола снижает общую производительность популяции, так как сокращает число особей, приносящих потомство. Это делает понятным, почему даже среди высших животных изредка наблюдаются партеногенетическне популяции. Наблюдения И. С. Даревского [Даревский, 1964; Darewski, Kulicowa, 1961], подробно изучившего экологию партеногенетических популяций скальных ящериц (Lacerta saxicola), делают это совершенно очевидным.
Однако в целом партеногенез не нашел широкого распространения среди высших форм жизни. Причины этого хорошо известны. Только перекрестное оплодотворение создает предпосылки для формирования многообразных генетических вариантов, на основе которых формируются оптимальные в данных условиях генотипы, и сводит к минимуму возможность рождения нежизнеспособных организмов 1. Однако этот же процесс вполне мог бы быть обеспечен гермафродитной популяцией, состоящей из особей, не способных к самооплодотворению. При этом сохранились бы все преимущества полового размножения, а потенциальная продуктивность популяции повысилась бы вдвое. Более того, гермафродитизм сни-
1 В настоящее время этот вопрос кажется более сложным, чем в недавнем прошлом. В популяции, размножающейся бесполым путем, две полезные мутации могут закрепиться лишь в том случае, когда одна из них возникает среди потомков ранее мутировавших особей. Среди организмов, размножающихся половым путем, обе мутации могут объединиться в результате рекомбинаций. Математический анализ показал [Crow, Kimura, 1965], что половой процесс выгоден в тех случаях, когда совместное действие мутаций усиливает полезный эффект каждой из них, когда мутирование происходит с высокой скоростью, а размеры популяции велики. С другой стороны, в тех случаях, когда индивидуальное действие мутаций отрицательно, а совместное положительно, рекомбинации могут оказаться вредными. Цитированные авторы сомневаются в безусловной полезности полового процесса и допускают возможность, что диплоидность следует рассматривать как механизм защиты от соматических мутаций. Общий вывод авторов подтверждается математическими расчетами Томлинсона [Tomlinson, 1966], который показал, что, когда малое число особей разбросано по большой территории, партеногенез и гермафродитизм выгодны. Если сомнения Кроу и Кимуры содержат в себе зерно истины, то это лишь с особой силой подчеркивает необходимость попытаться объяснить, в чем причина сексуализации всего живого мира.
{163}
мает и ряд трудностей, связанных со встречей брачных партнеров, возникающих в малочисленных популяциях. Тем не менее природа пошла по иному пути — по пути разделения полов. Единственное объяснение этой поистине удивительной расточительности природы заключается в том, что разделение вида на две генетические различные группы (самцы и самки) с неизбежностью ведет к возникновению и физиологических различий 1. Это в свою очередь с той же неизбежностью ведет к экологическим различиям, являющимся, как мы пытались показать, гарантией поддержания генетической разнородности популяции даже в крайне неблагоприятных условиях среды, сопровождающихся резкими снижениями численности. Отсюда следует, что значение поддержания генетической разнородности популяции настолько велико, что в конечном итоге компенсирует снижение потенциальной производительности популяций вдвое.
Генетическая разнородность популяций — предпосылка их эволюционных преобразований. Однако естественный отбор не может работать в кредит. Это значит, что генетическая разнородность популяций является не только предпосылкой их преобразований, но и повышает жизнестойкость популяции в текущий момент ее истории. Справедливость развиваемой точки зрения подчеркивается явлениями, сопутствующими сексуализации, на которые мы обращали внимание.
Мы приходим к выводу, что первопричиной разделения полов явилась необходимость поддержать всеми доступными способами максимальную разнородность популяции. На этой основе позднее возникли специальные анатомо-физиологические приспособления, завершающие разделение функций между самцами и самками, которое наибольшего развития достигает у млекопитающих. Ведущие механизмы поддержания генетической разнородности популяций — экологические. В преобладающем числе случаев они исключают возможность потери популяцией эволюционной пластичности и обеспечивают возможность быстрых приспособительных изменений генетической структуры популяции при изменении условий среды, изменении направления отбора. Поэтому экологические механизмы поддержания генетической разнородности популяций играют, вероятно, не менее существенную роль в эволюции вида, чем механизмы, обеспечивающие непосредственное преобразование популяций.
1 Весьма показательно, что у высших животных физиологические различия между полами неизмеримо более существенны, чем у низших.
{164}
Глава VI
ЭКОЛОГИЧЕСКИЕ МЕХАНИЗМЫ ПРЕОБРАЗОВАНИЯ ГЕНЕТИЧЕСКОЙ СТРУКТУРЫ ПОПУЛЯЦИЙ
Главная задача эволюционной экологии заключается в том, чтобы установить, как влияет популяционная структура вида на ход эволюционного процесса. Эта задача может быть сформулирована и иначе, более конкретно: какова взаимосвязь между экологической и генетической структурой популяции, как отражается изменение экологической структуры популяции на ее генетическом составе. В процессе решения этой задачи экологические механизмы микроэволюции могут быть вскрыты с наибольшей полнотой.
РОЛЬ ДИНАМИКИ ВОЗРАСТНОЙ СТРУКТУРЫ ПОПУЛЯЦИИ В ПРЕОБРАЗОВАНИИ ЕЕ ГЕНЕТИЧЕСКОГО СОСТАВА
Поддержание оптимальной возрастной структуры популяций — один из основных механизмов приспособления животных к конкретным условиям среды их обитания. Различные стороны этого вопроса в настоящее время интенсивно изучаются, им посвящена огромная литература. Однако изучение возрастной структуры популяций имеет не меньший интерес и для познания механизмов эволюционного процесса. Эта проблема изучена в значительно меньшей степени.
Сезонные изменения морфофизиологических особенностей долгоживущих животных (жизненный цикл которых охватывает по крайней мере несколько лет) хорошо изучены. Изменения теплоизоляционных свойств покровов, уровня энергетического обмена, количества и химического состава запасных питательных веществ и витаминов, потребности в кормах, деятельности важнейших органов эндокринной системы, общих (в том числе и поведенческих) реакций на изменение условий среды — вот далеко не полный перечень тех существенных физиологических сдвигов, которые легко обнаруживаются при сравнении животных на разных стадиях сезонного цикла их жизнедеятельности.
То же самое обнаруживается и при изучении мелких животных с коротким жизненным циклом. Однако в этом случае изменения свидетельствуют не только об изменении физиологических особенностей животных в процессе их развития, но и о морфофизиологической специфике животных разных поколений (генераций). Мелкие грызуны, насекомоядные и некоторые другие, хуже изучен-
{165}
ные группы животных, в полном смысле слова — эфемеры. У преобладающего числа видов этих групп перезимовавшие особи весной дают потомство, а к середине лета отмирают. Осенняя популяция состоит уже из других животных, родившихся во второй половине лета (то, что отдельные особи живут дольше основной их массы и доживают до одного года, не меняет общей картины). Осенняя популяция в высшей степени специфична. Животные этой генерации обладают еще более глубоким комплексом морфофизиологических особенностей, чем осенние животные долгоживущих видов. Это и понятно, ибо они с момента рождения развиваются в своеобразных условиях среды и выполняют вполне определенную экологическую функцию: они должны пережить зиму, весной дать потомство и передать эстафету жизни следующим поколениям. Нет возможности даже перечислить все те особенности, которые отличают осенние популяции, например полевок, от весенних. В том, пожалуй, и нет необходимости, так как материалы этого рода широко публикуются. Результаты, полученные по этому вопросу в нашей лаборатории, недавно сведены в коллективной статье [Шварц и др., 1964]. Смена биологически специфичных генераций обеспечивает более полное приспособление популяции в целом к сезонным изменениям условий среды, чем это возможно у видов, обладающих большой продолжительностью жизни, Представленные схемы (рис. 12, 13) показывают, какой сложности достигает смена сезонных генераций у грызунов в различных ландшафтных зонах.
Мы не всегда знаем, какие конкретные причины определяют морфофизиологические особенности конкретных генераций грызунов, но мы точно знаем, что все они могут быть сведены к двум принципиально различным факторам.
1. Морфофизиологическая специфика сезонных генераций — результат прямой реакции организма на изменение условий среды. Частное проявление этой закономерности — влияние физиологического состояния матери на организм потомства (исследования ряда лабораторий, в том числе и нашей, позволяют предположить, что некоторые физиологические особенности осенних генераций могут быть объяснены особенностями их матерей — животных, родившихся ранней весной).
2. Морфофизиологическая специфика сезонных генераций — это результат перестройки генетической структуры популяций, связанной с изменением направления отбора на разных стадиях жизненного цикла вида.
Значение первого фактора не вызывает сомнения. Оно доказывается большой серией экспериментов, показывающих, что изменяя условия существования можно вызвать «имитацию» сезонных изменений морфофизиологических особенностей вида в любое время года. Следует, однако, отметить, что подобные «несвоевременные изменения» обычно бывают все-таки менее резко выражены, чем истинно сезонные.
{166}
<![if !vml]>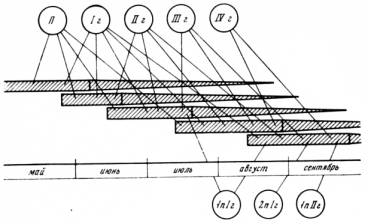 <![endif]>
<![endif]>
Рис. 12. Схема возрастной структуры популяции полевки-экономки (Microtus oeconomus chachlovi) на Крайнем Севере П — перезимовавшие особи; 1г—IVг — первая — четвертая генерации от перезимовавших; 1пIг—2п1г — первое и второе поколение особей I генерации; 1пIIг — первое поколение особей II генерации
Установив большую роль непосредственного влияния условий существования в формировании специфики сезонных генераций, мы не можем считать нашу задачу выполненной, так как нет сомнений в том, что в любых природных ситуациях конкретные особенности животного определяются не только условиями среды, но и особенностями генотипа. То, что особенности сезонных генераций отражают конкретные условия их развития, не исключает возможности параллельной перестройки генетической структуры популяций.
Доказано, что сезонная цикличность жизнедеятельности популяции в целом может сопровождаться изменением ее генетической структуры. Естественно, что наиболее отчетливо эта закономерность могла быть продемонстрирована па полиморфных популяциях. На представителях различных классов животных разными авторами было убедительно показано, что от весны к осени и от осени к весне генетическая структура популяции меняется: особи, лучше переносящие зимовку, преобладают весной, более плодовитые животные — осенью. Естественно, что преобладание в популяции определенных генетических вариантов меняется не только по сезонам, но и по годам («хронографическая изменчивость» — [Шварц, 1963]). С. В. Кириков [1934], кажется, впервые выяснил, что «черный хомяк» (черная фаза хомяка обыкновенного — Cricetus cricetus) распространен преимущественно в горных районах Башкирии и на северном пределе европейской части ареала вида. Автор считает вероятным, что черный хомяк более приспособлен к холодному и сырому климату. Позднее С. М. Гершензон [1945]
{167}
<![if !vml]>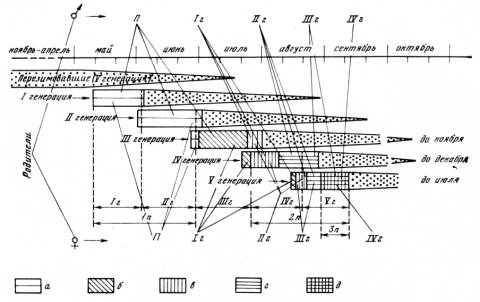 <![endif]>
<![endif]>
Рис. 13. Схема возрастной структуры и происхождения генераций полевых мышей (Apodemns agrarius)
П — перезимовавшие особи; 1г—IVг — первая — четвертая генерации от перезимовавших особей; In — 3п—потомство самок: а — перезимовавших, б — первой, в — второй, г — третьей, д — четвертой генераций
{168}
установил, что меланистические хомяки к осени достигают большей численности, а зимой они гибнут в большом числе. В. Н. Павлинин (устное сообщение) собрал интересный материал о встречаемости различных фаз хомяка по данным заготовок шкурок Сарапульской пушной базой. Проведенный нами анализ этого материала позволяет сделать некоторые интересные выводы. В отдельные годы соотношение черной и пестрой фазы изменяется очень резко. Так, например, в марте 1953 г. в Челябинской области на 253 пестрых хомяка приходилось 74 черных, а в декабре на 77 пестрых — 185 черных. В Башкирии, где вообще преобладают черные хомяки, летом в отдельные годы численность пестрых превышает численность черных, а к осени последние вновь начинают доминировать.
Фактов подобного рода можно было бы привести много, но еще более интересны другие, показывающие, что соотношение фаз меняется не только по сезонам, но и по годам. Осенью 1952 г. на территории Башкирской АССР было добыто 1357 пестрых хомяков и 1724 (56%) черных, а осенью 1951 г., соответственно 7248 и 3640 (34%). Т. В. Дмитриева [1949] показала, что соотношение серых длиннохвостых и желтых короткохвостых домовых мышей изменяется по годам. Она полагает, что серая «фаза» более восприимчива к туляремии и к неблагоприятным климатическим условиям. Согласно нашим наблюдениям, относительная многочисленность маланистов среди водяных крыс в лесостепном Зауралье меняется из года в год. В отдельные годы они составляют большинство в популяции. Почти по всему ареалу оппосума Trichosurus vulpecula встречаются меланисты, но они малочисленны, за исключением тасманийской популяции, в которой черная фаза преобладает. Пирсон [Pearson, 1938] установил, что меланисты лучше переносят холод и повышенную влажность. На моллюсках (Cepaea hortensis, С. nemoralis) показано [Schnetter, 1950; La Mot-te, 1959], что разные цветовые варианты обладают различной чувствительностью к изменению влажности. Физиологические различия голубых и зеленых гусениц некоторых бабочек определяют их различную чувствительность к ядам [McEwen, Splittstosser, 1964]. Аналогичными причинами объясняется различная смертность черных и полосатых тритонов в разных условиях [Test, 1954].
Среди амфибий диморфным видом можно считать озерную лягушку (R. ridibunda), но проявляется полиморфизм у этого вида только в молодом возрасте. Только что закончившие превращение лягушата четко разделяются на две группы: у одних спинка окрашена однотонно, у других вдоль хребта проходит узкая белая полоска (forma striata). Четкий диморфизм наблюдается и в пределах изолированных популяций на маленьких водоемах, что дает основание полагать, что он свойствен и особям, происходящим от одних родителей. Нами изучены некоторые интерьерные показатели у молодых озерных лягушек из Степного района Актюбин-
{169}
ской области. Оказалось, что две «фазы» лягушек заметно отличаются по такому важному признаку, как относительные размеры печени. У одновозрастных лягушат весом 2,2–3,0 г относительный вес печени равен в среднем: у лягушек с полосой— 50,2%о, у лягушек без полосы — 43,0.
Эйзентраут [Eisentraut, 1929] установил, что меланисты Lacerta lilfordi отличаются относительно более длинным кишечником. Автор высказывает предположение, что благодаря этому они обладают большей способностью использовать растительные корма. Естественно, что изменение условий среды вызывает изменение численного соотношения биологически различных фаз. Сезонные изменения генетической структуры популяций отмечались и другими авторами: Н. П. Дубининым и Г. Г. Тиняковым [1947] — на Drosophila funebris, Райтом и Добжанским [Wright, Dobzhansky, 1946] — на D. pseudoobscura, E. И. Лукиным [1962, 1966] — на пирокорисе, Н. В. Тимофеевым-Ресовским [Тимофеев-Ресовский, Свирежев, 1966] — на Adalia bipunctata и др. Подводя теоретический итог исследованиям этого направления, Н. В. Тимофеев-Ресовский [1964] писал: «Необходимой основой любой формы полиморфизма является длительное состояние динамического отборного равновесия между двумя или несколькими генотипами. Такое равновесие в свою очередь всегда основано на разном и конкурирующем давлении отбора трех существующих в популяции мутантных форм одного и того же гена или хромосомы (гетерозигота и две разные гомозиготы); или же конкурентные и разнонаправленные давления отбора двух или нескольких разных генотипов (из общей гетерогенной массы индивидов популяции) в различных (в пространстве или во времени) микроусловиях, наличествующих в пределах территории, занятой популяцией». Таким образом, изучение полиморфизма показало, что изменение условий существования и соответствующее изменение направления отбора приводят к изменению генетического состава популяции. В одни сезоны года преобладают одни генотипы, в другие — другие. Спрашивается, исключительное ли это явление, свойственно ли оно только явно полиморфным популяциям, в которых генетические различия между отдельными особями проявляются особенно резко, или оно свойственно любым популяциям и не обнаруживается лишь потому, что его технически трудно обнаружить?
Таким образом, первая половина нашей задачи заключается в том, чтобы доказать сам факт генетической перестройки популяции как закономерного явления. При этом особое значение имеют теоретические доказательства, так как даже десятки примеров в принципе не исключают возможности того, что наблюдаемое явление — уникальное.
Система наших доказательств сводится к следующему. Изучение полиморфных популяций показало, что сезонные изменения условий существования действительно связаны с изменением направления отбора. Это первая посылка. С другой стороны, дока-
{170}
зано, что генетическая разнородность популяции охватывает любые признаки организма, в том числе и такие, как плодовитость, скорость полового созревания, скорость роста, использование различных питательных веществ и т. п., значения которых в разные сезоны года резко различны. Так, например, Е. М. Масленникова и Д. Б. Хромач [1954] указывают, что среди крыс наблюдается сильно выраженная индивидуальная изменчивость потребности в витамине B2. Нетрудно допустить, что в условиях недостатка пищи, содержащей этот витамин, преимущество получают особи, потребность которых в указанном витамине выражена менее остро. Известны также индивидуальные варианты потребности в витамине Д, их наследственная закрепленность доказана [Harris, 1954]. В популяциях птиц отмечена генетическая изменчивость в отношении способности к использованию тиамина [Howes, Hutt, 1956].
Точными экспериментами доказано, что активность холинэстеразы в сенсорных областях коры у крыс подвержена индивидуальной изменчивости: крысы, обладающие большей активностью фермента, обладают более четкой реакцией на освещенность [Krech et al., 1954]. У различных генетических вариантов домовой мыши реакция молочных желез на эстрон и прогестерон оказалась различной [Mixner, Turner, 1957]; различна и восприимчивость тканей к гормону роста гипофиза [King, 1965]. Более того, даже такие признаки, как выраженность полового диморфизма [Korkman, 1957], скорость полового созревания в конце сезона размножения [Покровский, 1962], предпочитаемостъ разных кормов, выбор местообитания [Wecker, 1964], осторожность [Crowcroft, 1961], радиорезистентность [Bartlett et al., 1966], интенсивность биосинтеза гормонов [Badr, Spickett, 1965], подвержены индивидуальной изменчивости, детерминированной генетически.
Неоднородность популяции — биологический закон, не знающий исключений. Ему подчиняются любые признаки любых организмов. Поэтому изменение направления отбора неизбежно вызовет изменение генетической структуры популяции и каждая генерация становится специфичной не только физиологически, но и генетически. Изменение возрастной структуры популяции приводит, следовательно, к изменению ее генетической структуры.
Для примера воспользуемся наиболее стабильными признаками — краниологическими, фиксированными наследственностью в относительно узких рамках изменчивости. В последние годы стало ясным, что наиболее точно краниологические особенности животных характеризуют не абсолютные значения отдельных признаков и даже не их пропорции, а характер зависимости между общими размерами черепа и отдельных его частей. Вышла серия работ, демонстрирующая возможность использования аллометрии в таксономических целях [Huckinghaus, 1965; Rohrs, 1961]. Было показано, что аллометрический показатель (а) фиксирован на-
{171}
следственностью в значительно более узких рамках изменчивости, чем абсолютные размеры органов или частей тела, и практически не изменяется при изменении условий среды. Фрик [Frick, 1961] разделил колонию белых мышей на 2 группы. Одна из них развивалась в условиях, требующих резкого повышения физической нагрузки, другая служила контролем. Как и следовало ожидать, у подопытных мышей размеры сердца и почек значительно увеличились, но характер изменения размеров органов при изменении размеров тела остался неизменным. Аналогичные результаты были получены В. Г. Ищенко [1966, 1967] при сравнении размеров сердца полевки-экономки из природных популяций и лабораторной колонии (рис.14).
<![if !vml]>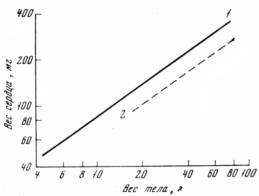 <![endif]>
<![endif]>
Рис. 14. Аллометрический рост сердца полевок-экономок (Microtus oeconomus oeconomus) популяции оз. Сасыкуль в Северном Казахстане (1) и в виварии (2)
В лабораторных условиях размеры органов уменьшаются (результат снижения уровня обмена), но а не изменяется. Эти и некоторые другие работы показывают, что характер соотносительного роста, выраженный в а, может служить очень хорошим показателем генетических отличий между популяциями и может быть использован при изучении динамики генетической структуры популяций.
Посмотрим, остается ли аллометрический экспонент постоянным в процессе смены сезонных генераций грызунов. Для этого воспользуемся материалами, любезно предоставленными лабораторией В. В. Кучерука (обработка материала В. Г. Ищенко). Предоставленный материал — это великолепная серия черепов, собранная в Волго-Ахтубинской пойме в разные годы и в разное время. Полученные материалы представлены в табл. 17. Они показывают, что в пределах популяции изменчивость а значительно превосходит межвидовые различия. Для того чтобы убедиться в этом, достаточно сопоставить данные табл. 17 и 18. Хотя это обстоятельство и не имеет прямого отношения к нашей теме, мы обращаем на него внимание. Еще раз показано, что без учета внутрипопуляционной изменчивости оценить различия между популяциями и даже видами часто бывает невозможно.
<![if !supportEmptyParas]> <![endif]>