
Проблемы Эволюции |
Из книги "Эволюция териодонтов". М.: Наука, 1976.
Фтинозухии.
Долгое время считалось, что наиболее примитивную группу терапсид составляют дейноцефалы, непосредственно связующие териодонтов и аномодонтов с пеликозаврами (Broom, 1914; Watson, 1914a, 1917 а, 1942), хотя ранее Сили (Seeley, 1894 ^ отметил особую примитивность "пеитерозавров" медистых песчаников Приуралья по сравнению с типичными южноафриканскими 'аномодонтами". Такие представления удерживались до последнего времени (Ефремов, 1954; Орлов, 1958; Huene, 1956), и, пожалуй, только Брум ( Broom, 1932) отметил, что наиболее древние тероцефалы морфологически примитивнее дейноцефалов.
Изменение взглядов на положение дейноцефалов в системе связано с открытием в нижних фаунистических горизонтах верхней перми СССР фтинозухий. Phthinosaurus первоначально относился Ефремовым к сфенакодонтным пеликозаврам, а позднее вместе с переизученным к тому времени Phthinosucchus - к горгонопсам (Ефремов, 1954), но уже. Ромер (Romer,1961) выделил их в особый подотряд Phchinosuchia, предковый как для настоящих териодонтов, так и для дейноцефалов. К аналогичному заключению пришел и Бунстра (Boonstra, 196За), объединивший фтинозухов с очерскими "эотериодонтами* в подотряде Eotitanosuchia.
Решение вопроса о положении в системе фтинозухий затрудняется значительным сходством всех примитивных терапсид, даже относящихся к различным эволюционным стволам. Кроме того, фтинозухий известны по единичным формам, существовавшим одновременно с дейноцефалами и относящимися, по-видимому, к различным концевым ветвям этой группы терапсид. Все же накопленный к настоящему времени материал определенно подтверждает взгляды Ромера— Бунстры о положении фтинозухий в основании родословного древа терапсид. Признаки особой примитивности фтинозухий, по сравнению с дейноцефалами, сближают этих териодонтов со сфенакодонтными пеликозаврами. К ним относятся; 1) общая конфигурация черепа с высокой и узкой мордой, плавно спускающимися вниз и назад заглазничным отделом и положением челюстного сочленения у заднего края черепа, примерно на уровне затылочного; 2) примитивное хищное озубление с узкими сжатыми с боков клыками, коническими резцами и многочисленными заклыковыми зубами; 3) образование заглазничной костью крыши над височной ямой без резко выраженных признаков прикрепления челюстных аддукторов к наружной поверхности этой кости у задневерхнего края надглазничной дуги; 4) узкие и практически не расширенные кпереди сошники; 5) вероятное сохранение, по крайней мере отдельными формами ( Eotitanosuchus), подвижного базиптеригоидного сочленения.
В отличие от Ромера и Бунстры мы считаем, что родственные связи фтино— зухий с горгонопсами, а отчасти и 'эвтериодонтами* были более тесными, чем с дейноцефалами.. Это оправдывает объединение их с типичными териодонтами в одном таксоне. Из числа признаков, сближающих фтинозухий с горгонопсами и "эвтериодонтами", следует отметить: 1) образование на небе хоанной ямы с некоторым погружением в нее сошников; 2) образование у эотитанозухид и биармозухид базикраниальных пластин крьшоввдных косгей с заключенной между ними" межптеригоидной ямой; 3) некоторое удлинение лицевого отростка септомаксиллы; 4) образование у Phthinosaurus зачаточного венечного огростка зубной кости.
Сфенакодонтные пеликозавры, фгинозухии и более поздние "типичные" териодонты представляют собой различные ступени одного направления специализации, тогда как дейноцефалы являются явно боковой ветвью с тенденцией перехода к растительному питанию.
Известные фтинозухии слишком специализированы, чтобы быть предками каких-либо других терапсид ( Boonstra, 1963a,b). В их строении сочетаются архаичные признаки с отдельными дейноцефаловыми и горгонопсовыми (Phthinosuchus) или же "эвтериодоновыми" и горгонопсовыми (Eotitanosuchidae, Biarmosuchidae). Так, фтинозух обладает практически горгонопсовым (но с парным сошником) нёбом и дейноцефаловым базиптеригоидным сочленением, а эотитанозухиды и биармозухиды - межптеригоидной ямой, близкой к эвтериодонтово-му типу. В то же время в форме и положении височного окна, а также в резко опущенном челюстном сочленении биармозухид проявляется некоторое сходство их с примитивными горгонопсами типа иктидоринид и пробурнетии (Татаринов, 1968). Совершенно уникально строение нижней челюсти у Phthinosaurиs, в которой сочетаются отдельные эвтериодонтные (скалопозавровые) и, возможно, офиакодонтовые признаки. К числу последних относятся: 1) исключительно высоко расположенная (на уровне верхнего края челюсти) сочленовная кость; 2) наличие окна под основанием- венечного отростка зубной кости, ведущего с наружной поверхности челюсти в аддукторную яму; 3) изогнутость верхнего края челюсти.
На родство со скалопозаврами (иктидозухами) указывает наличие у фтино-завра мандибулярного окна, а также отсутствие, по крайней мере у одного из скалопозавров (Scalopogaathus), выраженного, венечного отростка.
Горгонопсы
представляют специализированную примитивную ветвь, происходящую, по всей вероятности, от фтинозухий. Из числа характерных для последних и горгонопсов, но отсутствующих у типичных "эвтериодонтов" признаков отметим: 1) широкую крышу над височными ямами, образованную заглазнич-ными костями; 2) сильно развитые нёбно- птеригоидные озубленные бугры и зубы на поперечных флангах крьшоввдных костей; 3) тропибазальную мозговую коробку с полно окостеневающим сфенэтмоидом. С фтинозухиями горгонопсов сближает также отсутствие подглазничных окон и подвижное (по крайней мере, у типичных представителей) базиптеригоидное сочленение. К числу признаков собственной специализации горгонопсов относятся наличие на крыше черепа предтеменной кости и обычное образование глубокой хоянной ямы.
Целесообразность противопоставления горгонопсов "эвтериодонтам" оправдывает, однако, не столько наличие у них большого числа примитивных особенностей, сколько отсутствие прямых родственных связей между горгонопсами и какими-либо эвтериодонтами. Долгое время широким признанием пользовались взгляды Уотсона (Watson,1914 b,1921, 1942), согласно которым горгонрп-. сы считались прямыми предками цинодонтов. Однако такое предположение основывалось главным образом на поверхностном сходстве обеих групп в строении нёба (отсутствие подглазничных окон и эвтериодонтовая межптеригоидная яма). С открытием (Broom, 1938) примитивнейших цинодонтов - процинозухов, сохранявших хорошо "развитую межптеригоидную яму, эти аргументы утратили значение.
Сигоньо и Чудинов ( Sigogneau, Tchudinov, 1972) склонны считать, что наибольшей близостью к горгонопсам обладают биармозухиды. Мозаичное проявление горгонопсовых особенностей среди трех семейств фтинозухии - фтинозухид, биармозухид и эотитанозухид - делает такое утверждение преждевременным; каждое из перечисленных семейств, кроме того, имеет и признаки собственной специализации. Наиболее близки к горгонопсам, по нашему мнению, фтинозухиды, при этом горгонопсы ответвились от "эотериодонтов" на несколько более примитивном уровне.
Среди горгонопсов наиболее примитивное положение занимает сем. Ictidorhinidae (Sigogneau, 1963a, 1968,1970). обладающее значительным сходством в пропорциях с некоторыми эотериодонтами типа Biarmosuchus. Боковое ответвление примитивных горгонопсов представляет сем. Burneciidae (Татаринов, 1968). В строении обеих семейств отмечаются черты сходства с дейноце-фалами, имеющие, по-видимому, конвергентную природу. У иктидоринид это сходство выражается в 'перекресте" резцов (Sigogneau, 1968, 1971), а у бурнетиид - в резком утолщении костей черепа. Отсутствие и у иктидоринид, и у бурнетиид венечного отростка придает им дополнительное сходство с фтинозу-хиями. Оба эти семейства почти с одинаковым основанием могут относиться как к фтинозухиям, так и к горгонопсам: грани между ними выражены неотчетливо. Поэтому нам кажется целесообразным объединение обеих групп в одном отряде - Gorgonopiain sensu latum, который может подразделяться на подотряды Phthinosuchia и Gorgonopsia in sensu scrictum (Eugorgonopia).. Сем. Phthinosauridae занимает среди фтинозухий обособленное положение. Центральную группу эвгоргонопсов образуют Gorgonopidae, приобретающие отдельные ''териодонтовые" признаки (удлиненные базикраниальные пластины птеригоидов} независимо от 'эвтериодонтов*. Сем. Inoscranceviidae и Rubidgeidae можно рассматривать в качестве параллельно специализированных ответвлений горгонопид, самостоятельно выработавших качества гигантского саблезубого хищника. Broomicephalinae занимают среди рубиджиид несколько обособленное положение (Sigogneau, 1970).
Эвтериодонты.
Под этим названием мы выделили в особый отряд всех высших териодонтов, морфологически более далеких от фтинозухий, чем собственно горгонопсы (Татаринов, 1974). Накапливающийся фактический материал приводит, однако, к выводу о целесообразности разделения эвтериодонтов на два отряда: Therocephalia in sensu latum и Cynodontia, каждый из которых, по-видимому, происходит от фтинозухий самостоятельным стволом.
Тероцефалы.
В этом отряде мы объединяем подотряды мигалезавров (Mygalesaura), собственно тероцефалов (Therocephalia in sensu strictum, seu Scylacosauria) и скалопозавров (Scaloposauria). Общим для всех тероцефалов является наличие типичных подглазничных окон, утрачиваемых лишь вайтсиидами и, возможно, некоторыми скалопоцинодонтами.
Мигалезавры
занимают среди тероцефалов совершенно изолированное положение, отличаясь, в частности, наличием широкой крыши над височными окнами, образованной заглазничными костями. Этот признак придает им определенное сходство с фтинозухиями и собственно горгонопсами, и если бы не формирование типичных подглазничных окон, мигалезавров можно было относить к фтинозухиями. Однако позднее появление мигалезавров в геологической летописи (они известны только из зоны Kiscecephalus Южной Африки; и усложнение заклыковых зубов, имеющих трехзубчатое строение, заставляют предполагать длительную и неизвестную пока эволюционную историю мигалезавров. Поэтому мы предпочитаем рассматривать мигалезавров в качестве особого подотряда, а не присоединять их к фтинозухиям, как предложили Кун (Kuhn,1966) и Ромер (Romer,1966a, 1971).
Сцилякозавры
морфологически в общем примитивнее скалопозавров и цинодонтов, однако сколько-нибудь явных родственных связей с высшими териодонтами они не имеют, представляя собой по существу слепую ветвь. Предположение Кемпа (Кетр,1972а) о происхождении цинодонтов от тероцефалов вайтсиидного типа недостаточно обосновано (см. ниже).
В геологической летописи сцилякозавры появляются в основании зоны Tapinocephalus Южной Африки и нижнетатарском подъярусе Восточной Европы. Разрыв между эветроцефалами и известными фтинозухиями велик и происходить они могли только от еще неизвестных, малоспециализированных и, по-видимому, мелких представителей последних. Наличие эвтериодонтовой межптеригоидной ямы и предклыковых зубов в челюстной кости заставляет предполагать лишь очень отдаленные родственные связи примитивных сцилякоаавров с фтинозухиями, возможно, эотитанозухидного облика. По общей конфигурации черепа с резко вынесенным вниз челюстным сочленением и некоторым изгибом базикраниальной оси больше отличаются от сцилякозавров биармоаухиды.
Наиболее примитивную группу сципякозавров образует надсем. Scylacosauroidea. Среди них Priscerognathidae, имеющие массовое распространение в зоне Tapinocephalus Южной Африки, обычно рассматриваются в качестве исходного для сцилякозавров (Boonstra, 1954; Вьюшков, 1955; Huene, 1956; Watson, Romer, 1956). Однако пристерогнатиды явно специализированы по сравнению с Scylacosaundae, Они утратили, в частности, предклыковые зубы в челюстной кости, а крыша черепа в височной области имеет у них узкий гребень цинодонтового типа. Поэтому мы склонны рассматривать птистерогнатид в качестве бокового ответвления от общего ствола сцилякозавроидей.
Морфологически к пристерогнатидам очень близко сем. Lycosauridae, характеризующееся, в частности, сходной конфигурацией черепа, необычно высокого для тероцефалов. Расширенность восходящей пластины эпиптеригоидов, однако, сближает ликозаврид с высшими тероцефапами иадсем. Whaitsioidea. По-видимому, ликозавриды приобрели расширенные эпиптеригоиды независимо от вайтсиоидей.
Тесные родственные связи между семействами вайтсиоидей — Annatherapsididae, Moschorhinidae, Euchambersiidae, Moschowhaitsiidae Whaksiidae - достаточно ясны. Эти эвтероцефалы отличаются, в частности, значительно уплощенным ("типично тероцефаловым"} черепом. Центральную группу среди перечисленных семейств образуют аннатерапсидиды, еще сохраняющие заднелобные кости. Интересно, что аннатерапсидиды обладают также зачаточными поперечными отростками на сошнике и челюстных костях, служившими основой для формирования вторичнонёбного костного мостика у мосховайтсиид и вайтсиид. Вторичное нёбо приобретено обоими названными семействами параллельно. В отличие от вторичного неба у бауриоморфов и цинодонтов оно выполняет исключительно механическую функцию - укрепление челюстного аппарата, но отнюдь не разделение дыхательного и пищеварительного трактов. Вайтсииды резко отличаются от аннатерапсидид также утратой подглазничных окон и заклы-ковых зубов. Мосховайтсииды сохраняют некоторые примитивные признаки, утраченные вайтсиидами — большие подглазничные окна и заклыковые зубы. Особый ствол, также исходящий от тероцефалов типа аннатерапсидид, образуют мосхориниды, отличающиеся резко укороченным черепом. Специализированная ветвь мосхоринид представлена эвчамберсиидами, утратившими все заклыковые зубы и приобретшими массивную связанную с клыком ядовитую железу. Формирование последней привело к образованию у Euchambersia обширной вырезки позади клыка, а также к утрате заглазничной и скуловой дуг (Nopcsa, 1933).
Скалопозавры в известной мере заполняют пробел между сцилякозаврами и цинодонтами. В геологической летописи примитивные скалопозавры (иктидозухи) появляются одновременно с сцилякозаврами. По общей конфигурации черепа с относительно широкой височной крышей, узкой и глубоко вырезанной сзади небной пластиной предчелюстной кости и сохранению предклыковых зубов в челюстной кости иктидооухи напоминают сцилякозавров сем. Scylacosauridae. Однако иктидозухи сближаются с высшими сцилякозаврами резким расширением восходящей пластины эпиптеригоидов и утратой заднелобных костей. По всей вероятности, оба признака были приобретены иктидозухами независимо от высших сцилякозавров. Во всяком случае отсутствие у абберантных нижнетриасовых иктидозухов сем. Scalopognathidae выраженного венечного отростка указывает на весьма раннее обособление скалопозавров от сцилякозавров. Не исключено даже, что ответвление скалопозавров произошло еще на фтинозуховом уровне и что отряд Therocephalia является филогенетически сборным. В числе дополнительных аргументов, свидетельствующих в пользу родства скалопозавров с фтиноаухиями, можно отметить наличие у эотитанозухид миниатюрной, но врезающейся в тело базисфеноида, как и у примитивных скало;— позавров, межптеригоидной ямы, а также развитие у phthinosaurus окна под венечным отростком нижней челюсти, хорошо выраженного и у многих иктидозухов.
Из числа иктидозухов изолированное положение помимо Scalopognathidae занимают Lycideopidae и Chthonosauridae, обычно рассматриваемые в составе сцилякозавров. Оба эти семейства отличаются от типичных иктидозухов крупными размерами, относительно высоким черепом, отсутствием сколько-нибудь выраженного вторичного неба. Уникально развитие на нёбных костях у Chthonosaurus радиальных, покрытых шагреневыми зубками гребней. Этот, по сути дела котилозавровый, признак ранее не отмечался у терапсид.
У высших иктидозухов (сем. Ictidosuchidae и Scaloposauridae) мы встречаемся с широким развитием параллелизмов, выражающихся в развитии зачаточного вторичного нёба, подразделении дистального конца парокципитального отростка, утрате заглазничной дуги, усложнении заклыковых зубов. Эти процессы крайне затрудняют решение вопросов родственных связей между высшими иктидозухами. Нам представляется преждевременной поэтому попытка Уотсона и Ромера (Watson, Romer,1956) разделить их на серию самостоятельных семейств.
Происхождение типичных триасовых скалопоэавров - бауриоморфов - от высших иктидозухов ( Watson, 1931) не может оспариваться. Однако более определенно высказаться о существующих между обеими группами родственных связах очень трудно. Бауриоморфов отличает от иктидозухов главным образом развитие полного вторичного нёба и усложенных заклыковых зубов. Эти признаки в менее выраженной форме проявляются у самых различных представителей Scaloposauridae и кажется вероятным, что каждое из трех семейств бауриоморфов - Nothogomphodoncidae, Ericiolacercidae и Bauriidae - происходит от скалопозаврид самостоятельно. В таком случае бауриоморфы представляют собой группу, выделенную по горизонтальному принципу, хотя и объединяющую тесно родственные семейства.
Для наиболее распространенных бауриоморфов - бауриид характерны развитие на заклыковых зубах широких уплощенных поверхностей и, вероятно, приспособление к растительному питанию (Crompton, 1962). Нотогомфодонтиды из среднего триаса Приуралья, обладавшие малоспециализированными трехзубчатыми заклыковыми зубами, питались, по-видимому, преимущественно животной пищей (Татаринов, 1974).
Наиболее проблематично положение в системе скалопоцинодонтов - небольшой группы малоизученных форм, отличающихся сложной мозаикой цинодонто— вых и иктидозуховых признаков и то относимых к цинодонтам (Haughton, Brink, 1954; Brink, 1961b; 1963с; Hopson, Citct»ing, 1972), то распределяемых между различными семействами иктидозухов ( Watson, Romer, 1956). По данным Бат— тейла ( Bactaile, личное сообщение), подглазничные окна на небе отсутствовали, по крайней мере, у одного из родов скалопоцинодонтов - у Silphedestes, на основании чего он категорически высказывается в пользу принадлежности скалопоцинодонтов к процинозухидным цинодонтам, с которыми их сближает и раз-деленность вторичного нёба щелью. По общей конфигурации черепа, однако, скалопоцинодонты близки к скалопозавридам. Кроме того, небольшое подглазничное окно имелось у Scalopocynodon,a хорошо развитое - у недавно открытого нижнетриасового скалопоцинодонта из Приурапья. Поэтому представляется возможным родство скалопоцинодонтов с иктидозухами; в таком случае подглазничные окна должны были быть утрачены скалопоцинодонтами вторично. Не исключена, однако, и гетерогенность скалопоцинодонтов, к которым, возможно, относят как родичей иктидозухов, так и процинозухов скалопозаврид-ного облика.
Цинодонты.
Поскольку Procynosuchidae обладают хорошо развитой межптери— гоидной ямой, Брум (Broom, 1938) на этом основании пришел к выводу о происхождении цинодонтов от тероцефалов. В дальнейшем Бринк ( Brink, 1961 в, 1963с) высказал взгляд о происхождении цинодонтов от иктидозухов через посредство скалопоцинодонтов, а Кемп ( Кетр, 1972а) - от высших тероцефалов вайтсиидного типа и, наконец, Ромер ( Romer, 1969a) уазал, что цинодонты, скорее всего, происходят от архаичных териодонтов фтинозухового типа.
Последнее предположение мы считаем наиболее убедительным. Гипотезу Бринка можно принять лишь в том случае, если признать, что все скалопоци-нодонты занимают переходное положение между иктидозухами и процинозухами. Сходство цинодонтов с вайтсиидными тероцефалами, акцентируемое Кемпом, проявляется главным образом в признаках, развивающихся параллельно в различных ветвях териодонтов (расширенность эпиптеригоидов, отсутствие заднелобной кости и пинеального отверстия. Более уникально отсутствие подглазничных окон, однако против родства цинодонтов с вайтсиидами говорит и различный для обеих групп способ формирования вторичного костного нёба, и конфигурация черепа, значительно более высокого у процинозухов, и большее число заклыковых зубов у цинодонтов, в чем они сходны только с фтинозухами. Между нёбными пластинами предчелюстных костей у цинодонтов постоянно развиты вырезки — нёбные щели, отсутствующие у вайтсиоидей, а нижняя челюсть намного массивнее, чем у тероцефалов. Наконец, межптеригоидная яма у прицинозухид намного крупнее, чем у тероцефалов, хотя она и не образует вырезки в теле базисфеноида, как у иктидозухов.
Цинодонты наиболее, пожалуй, интересная в теоретическом отношении группа териодонтов, отличающаяся распространением в ней "маммализации" - постепенного и параллельного в ряде ветвей приобретения признаков, характерных для млекопитающих. Исходное положение среди цинодонтов занимают процинозухии. Различия между ProcynosuchidaeHGalesauridae весьма условны. Среди галезаврид известны формы с неполным вторичным нёбом и сохраняющейся межптеригоидной ямой (Brink, 1965 b). Вполне возможно, что 'галезавридовый" уровень достигался различными представителями этого семейства параллельно. Наиболее обособленную группу процинозухий образуют Dviniidae, приобретшие полное вторичное нёбо и многобугорчатые заклыковые зубы, дифференцированные на предкоренные и коренные. В отличие от высших гомфодонтных цинодонтов, заклыковые зубы у двиниид не соприкасались при смыкании челюстей.
Высшие цинодонты - циногнатии приближаются к млекопитающим резко выраженной редукцией постдентальных костей нижней челюсти. Небольшая и, по-видимому, более примитивная группа циногнатии - Cynognachoidea - сохраняет примитивный хищный образ жизни. Основная группа ~ Gomphognachoidea перешла к растительному питанию и приобрела многобугорчатые : зубы со сложными жевательными поверхностями, широко соприкасающимися при смыкании челюстей.
Некоторые авторы предполагают, что редукция постдентальных костей проходила у хищных и растительноядных цинодонтов параллельно и объединяют на этом основании циногнатоидей в одной группе с процинозухиями (Hopson, Kitching, 1972).
Наиболее "маммалоподобные" цинодонты - тритилодонты и диартрогнаты -долгое время объединялись под названием иктидозавров и не включались в состав цинодонтов. Гетерогенность иктидозавров теперь можно считать доказанной (Crompton,1958). Тритилодонты происходят, несомненно, от грмфогна-тоидей, возможно, от форм, близких к предкам Traversodontidae или непосредственно от последних (Crompton, Ellenberger, 1957). Диартрогнаты некоторое время рассматривались в качестве потомков бауриоморфов ( Crompton,195 8). Однако типичные бауриоморфы явно слишком специализированы, чтобы быть предками диартрогнатов. Факты, послужившие основой для предположения о происхождении диартрогнатов от бауриоморфов - наличие у них обширной межптеригоидной ямы и сравнительно широкой теменной области крыши черепа, совершенно недостаточны для такого вывода. На основании этих же фактов допустимо происхождение диартрогнатов и от иктидозухов (Crompton, 196За), и от скалопоцинодонтов и, наконец, от процинозухов (В argbus en,1968). К последнему наиболее обоснованному заключению склоняется теперь и Кромптон (Crompton,1968aiCrompton, Jenkins, 1968). Отметим все же, что по размерам и форме межлтеригоидной ямы диартрогнаты наиболее близки к иктидозухам, а не к процинозухидам.
Диартрогнаты считались ранее уникальной группой териодонтов на основании наличия у них двойного челюстного сочленения. Теперь, однако, в той или иной степени выраженное дополнительное челюстное сочленение обнаружено у многих высших цинодонтов ( Crompton, 1927 а), а возможно, и у тритилодонтов ( Fourie, 1968). Формирование дополнительного челюстного сочленения, редукция постдентальных костей нижней челюсти и образование полного вторичного костного нёба осуществлялось у диартрогнатов независимо от остальных цинодонтов.
Таким образом, мы считаем возможным предложить следующий вариант классификации териодонтов:
Надотряд Theriodontia
Отряд Gorgonopia
Подотряд Phthinosuchia
Подотряд Eugorgonopia
Отряд Therocephalia
Подотряд Mygalesauria
Подотряд Scylacosauria
Подотряд Scaloposauria
Отряд Cynodonria
Подотряд Procynosuchia
Подотряд Cynognathia
Подотряд Tritylodontia
Подотряд Diarthrignathia
О параллелизмах в процессе маммализации териодонтов.
Общеизвестно, что высшие териодонты - диартрогнаты и тритилодонты - с трудом отличаются от примитивных мезозойских млекопитающих (Olson, 1944, 1959; Brink, 1957, 1967; Van Valen,1960; Reed, I960; Татаринов, 1965a, 1972a; Parrington, 1967). В качестве условного демаркационного признака между классами используют состав челюстного сочленения (Simpson, 1960), которое у млекопитающих образовано зубной и чешуйчатой, а у остальных тетрапод - сочленовной и квадратной костями. Однако среди териодонтов дополнительным челюстным сочленением обладали многие циногнатоидеи, гомфогнатоидеи, диартрогнаты и, возможно, некоторые тритилодонты ( Fourie, 1968; Crompton, 1972a), а среди мезозойских млекопитающих по крайней мере Morganucodon (Kermack,1965; Kermak ее al., 1973) обладал двойным челюстным сочленением.
Обычно считают, что процесс маммализации серьезное значение приобрел только у наиболее прогрессивных триасовых териодонтов. Однако уже самые древние из них, пермские, приобретали многие маммальные особенности, причем набор таких признаков и темпы их формирования оказываются различными для разных групп, о чем наглядно свидетельствуют некоторые примеры.
Вторичное костное небо, сделавшее возможным пережевывание пищи без приостановки дыхания, приобретено параллельно цинодонтами (двумя—тремя ветвями) и бауриоморфами. Придаточные бугорки на коронках заклыковых зубов появляются независимо у рубиджиний, скалопозавров и примитивных ци— нодонтов. Сложные жевательные коронки формируются параллельно в различных группах цинодонтов, у скалопогнатид и бауриоморфов. У северодвинских цинодонтов (двиниид} сложные жевательные коронки с многочисленными придаточными бугорками (более 20 на нижних зубах) сформировались еще до приобретения маммального прикуса со смыканием верхнего и нижнего рядов эаклыковых зубов.
Параллельно у циногнатий, тритилодонтов и диартрогнатов проходила и перестройка челюстной мускулатуры по маммальному образцу, приведшая к ослаблению нагрузки на челюстной сустав и усилению нагрузки на заднюю часть зубной кости. Этот процесс вел к разрастанию зубной кости, редукции постдентальных костей нижней челюсти и создал предпосылки для формирования маммального челюстного сочленения между зубной и чешуйчатой костями (Crompton,1963a,b;Barghusen, 1968, 1970, 1972; Barghusen, de Mar,1972; Mussett, 1967). По крайней мере частично постдентальные кости сохранялись у большинства мезозойских млекопитающих (Kiihne,1958; Kennack, Musset, 1958). У верхнетриасового Morganucodon сохранялось и рептилийное челюстное сочленение между сочленовной и квадратной костями, еще не преобразованными в слуховые косточки (Kermack, 1965). Сохранение рептилийного короной— да было установлено даже у иантотериев - ближайших предков современных плацентарных (Krebs, 1969).
Органы обоняния, резко усложненные у млекопитающих, во многих существенных деталях унаследованы ими от териодонтов. Обонятильные капсулы у последних резко увеличены и иногда составляли почти половину длины черепа. Уже у пермских териодонтов имелись хорошо развитые верхние обонятельные раковины маммального типа, о чем можно судить по наличию характерных парасагиттальных гребней на вентральной поверхности носовых костей.
"Протовибриссы" и зачаточные мягкие губы маммального типа также, видимо, были приобретены уже териодонтами. У пермских тероцефалов верхнегубные сосуды и нервы оставляют на костях развитую сеть отпечатков, сравнимую с сетью вибриссовых сосудов и нервов млекопитающих, у которых они проходят, однако, в толще хорошо развитых мягких губ. У наиболее прогрессивных териодонтов - у цинодонтов, тритилодонтов и, по-видимому, диартрогнатов — отпечатки сети сосудов и нервов исчезают с костей, сосудистые отверстия в челюстной кости объединяются друг с другом и отодвигаются далеко назад. Кроме того, у северодвинских цинодонтов имеются отпечатки губных мышц. Возможно, что у цинодонтов, как и у млекопитающих, формировались мягкиб губы и вибриссовые сосуды и нервы проходили в их толще. Приобретение мягких губ имело существенное значение в эволюции млекопитающих, будучи одной из предпосылок для формирования сосущего ротового аппарата и тем самым для перехода к вскармливанию молоди молоком.
Приобретали высшие териодонты и маммальные особенности кровоснабжения мозга, которое у млекопитающих заметно возрастает за счет участия наряду с внутренней сонной еще и подключичной артерии, в то время как из мозга кровь выносится не только средней и задней мозговыми венами, но и мощно развитой системой венозных выпускников. Особенно развита вена височного канала, иногда превышающего по диаметру яремное отверстие. Признаки развития маммальной системы венозных выпускников обнаружены у северодвинских цинодонтов. Крайняя узость сонных каналов у большинства териодонтов дает основания предполагать наличие у них дополнительного источника снабжения мозга артериальной кровью.
В различных группах териодонтов также независимо образуются видиевы каналы маммального типа, а базиптеригоидное сочленение утрачивает подвижность. Восходящая пластина эпиптеригоида расширяется, переднеушная кость образует сильно развитые отростки, подразделяющие надвое эпиптериковую полость, чем достигается состояние, исходное для мезозойских млекопитающих. Череп у триасовых териодонтов становится платибазальным, затылочный мыщелок парным, а конечности подворачиваются под тело, как и у млекопитающих. Большие полушария головного мозга резко расширяются у вайтсиоидей, бауриоморфов и цинодонтов.
Разобранных фактов явно достаточно для вывода о глубокой древности многих существенных маммальных признаков и о широком значении параллелизмов в процессе преобразования терйодонтовой организации в маммальную (см. рис. 41).
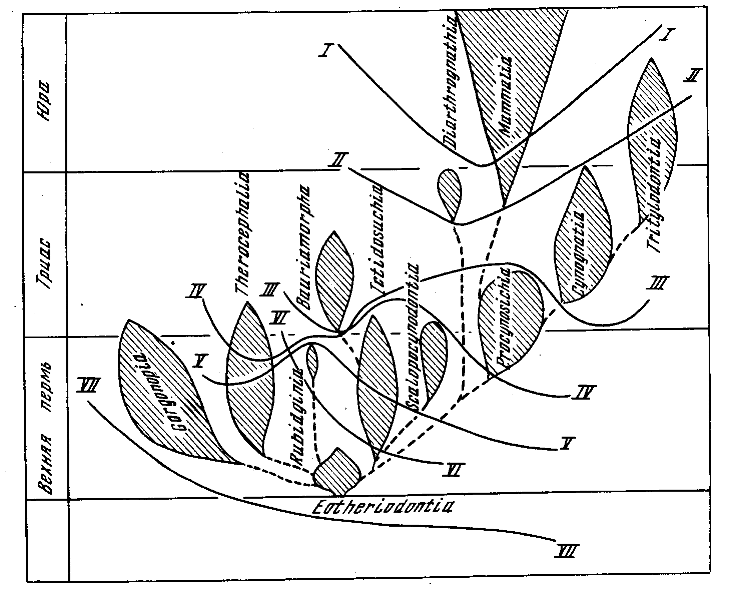
Рис. 41. Схема филогенеза и процесса маммализации териодонтов. Кривые линии соответствуют уровням, на которых приобретались: I - звукопроводящий аппарат из трех косточек; II - челюстное сочленение между зубной и чешуйчатой костью; 111 — мягкие, снабженные зачаточной мускулатурой губы; IV - расширенные большие полушария головного мозга; V - трехбугорчатые щечные зубы; VI - верхние обонятельные раковины (Татаринов, 1972)
О
происхождении млекопитающих. Первые годы после выхода в свет "Происхождения видов" проблема происхождения млекопитающих решалась главным образом трудами зоологов и сравнительных анатомов, путем сравнения рецент— ных млекопитающих с представителями низших классов наземных позвоночных. Геккель (Haeckel.1866) выводил млекопитающих из пресмыкающихся. Гексли(Huxley, 1871) видел предков млекопитающих в земноводных, обосновывая этотвзгляд платибазальностью черепа, парностью затылочного мыщелка, богатством кожи представителей обоих классов многоклеточными железами, а также сохранением у млекопитающих только левой дуги аорты (У птиц сохраняется только правая дуга аорты, у пресмыкающихся она получает преобладающее развитие, тогда как у земноводных в одинаковой степени развиты как правая ("птичья"), так и левая ("маммальная") дуги аорты)..
Взгляды Гексли произвели большое впечатление на современников, именно на них основано широко распространенное противопоставление пресмыкающихся и птиц (завропсид) млекопитающим. Тем не менее большинство немецких анатомов (Гегенбаур, Видерсгейм, Гаупп и др.) следовали за Геккелем, придававшим особое значение развитию у пресмыкающихся, птиц и млекопитающих амниона и противопоставившего все три класса амфибиям под названием Amniota.
Открытие зверообразных пресмыкающихся послужило началом палеонтологического исследования вопроса о предках млекопитающих. Уже Оуэн (Owen, 1856) отметил сходство в посткраниальном скелете между дицинодонтами и млекопитающими, но только Коп (Соре, 1878,1884) первым ясно высказал мнение о прямых родственных связях между зверообразными и млекопитающими, Последовавшее вслед за этим быстрое накопление данных по южноафриканским териодонтам дало мнению Копа серьезно аргументированную поддержку (Ваиг, Case, 1897). Некоторый диссонанс внесла вначале идея Сипи (Seeley, 1888), видевшего в териодонтах не прямых предков млекопитающих, а лишь группу, эволюировавшую параллельно с последними. Позднее Сили объединил южноафриканских зверообразных с клоачными млекопитающими в группу Theropsida, параллельную завропсидам, и .тем самым стал одним из основоположников взглядов о полифилетическом происхождении млекопитающих (Seeley, 1896, 1898). Однако после работ Осборна ( Osborn, 1888, 1898, 1903) теория происхождения млекопитающих от териодонтов завоевала общее признание. Дополнительные материалы, обработанные в последующие годы Брумом, дали этой теории богатейшую фактическую основу (Broom, 1905, 1914; Arldt, 1908). Тем не менее отдельные авторы до самого последнего времени видят в териодонтах лишь группу, параллельную млекопитающим, и считают предками последних гипотетических Sauromammalia (Haeckel, 1916) или Multiseriata (Матвеев, 1968).
Дальнейший сдвиг произошел лишь в 20-х годах нашего столетия в связи с расширением знаний о мезозойских млекопитающих. Мост, соединяющий пресмыкающихся с млекопитающими, стал наводиться с двух сторон. Однако контакта между палеотериологами и палеогерпетологами долгое время не было. До самых последних лет специалисты по териодонтам и мезозойским млекопитающим работали совершенно независимо и в результате сформулированные ими выводы оказались во многом различными.
Специалисты по териодонтам склонялись к идее монофилетического происхождения млекопитающих от высших териодонтов — "иктидозавров" (Broom, 1929, 1932; Watson, 1942, 1953; Crompton, 1958). В то же время специалисты по мезозойским млекопитающим с большим скепсисом относились к попыткам связать между собой различные отряды их, и в 1928 г. Симпсон заключил, что по крайней мере четыре группы млекопитающих произошли от териодонтов самостоятельно (Simpson, 1928). Такими параллельными ветвями являются, по Симпсону, клоачные, многобугорчатые, триконодонты и симметродонты. Последние, возможно, дали начало пантотериям, а вместе с ними и современным сумчатым и плацентарным. В последующем Симпсон стал допускать возможность самостоятельного происхождения от териодонтов еще докодонтов и пантотериев ( Simpson,1959). Авторитету Симпсона во многом обязано широкое распространение в эти годы полифилетической концепции происхождения млекопитающих (Bohlin,1945; Patterson, 1956; Kermack, Mussett, 1958; Kuhne, 1958;, Simpson, 1959, I960, 196la; Kermack, 19б5,19б7а,b).
Факты, добытые к этому времени специалистами по териодонтам, показали широкое распространение у зверообразных процессов параллельного формирования маммальных особенностей (Olson,1944, 1959; Brink, 1957, 1967; Татаринов, 1965а, 1970; Lessertisser, Sigogneau, 1967). Доказательства полифилии "иктидозавров" ( Crompton, 1958), распавшихся на тритилодонтов и диартрогнатов, еще более подорвали уверенность в монофилетическом происхождении млекопитающих. Полифилия млекопитающих стала большинством исследователей считаться серьезно обоснованной. Однако все предполагаемые родственные связи между отдельными группами мезозойских млекопитающих и териодонтами оставались неясными, почти произвольными. Один и тот же автор, подчас никак не аргументируя своих взглядов, резко менял точку зрения на этот вопрос. Так, Олсон (Olson) в 1944 г. выводил из иктидозавров только многобугорчатых млекопитающих, а в 1959 г. связывал с иктидозав-рами уже трикодонтов; докодонтов по непонятным причинам он выводил из бауриоморфов, а многобугорчатых - из тритилодонтов. Существенно менялись взгляды на родственные связи териодонтов с млекопитающими и у Бринка. Кюне в опубликованной почти одновременно с Олсоном работе, выводил докодонтов и большинство остальных млекопитающих из цинодонтов, а с бауриоморфами и иктвдозаврами сопоставлял только многобугорчагых (Kiihne,1958).
Сдвиг в решении этого вопроса наметился в результате попыток отдельных авторов, как палеотериологов, так и палеогерпетологов, возродить в той или иной форме прежнюю теорию эволюции зубов млекопитающих Копа - Осборна (Osborn, 1907). По этой теории, считавшейся совершенно отброшенной под влиянием работ Симпсона, коренные зубы различных млекопитающих выводятся из триконодонтовых посредством появления новых и смещения ('вращения") старых бугорков. Сначала Батлер (Butler, 1939) вновь попытался вывести сим-мтеродонтовые зубы из триконодонтовых, что означало уменьшение числа параллельно возникших ветвей млекопитающих на одну. Выводы Батлера, поддержанные Паррингтоном ( Parrington, 1947) и Паттерсоном (Patterson, 1956), привели ко всеобщему признанию новой концепции эволюции коренных зубов млекопитающих, в которой все их многообразие выводится уже не из протодонтного, а из триконодонтного зуба. В отличие от взглядов Копа и Осборна, гомологизировавших с главной вершиной рептилийного зуба протокой (в верхних) и протоконид (в нижних) коренных зубах, теперь принято мнение о вторичном развитии протокона. С главной вершиной рептилийного зуба сопоставляют теперь паракон и протоконид (Butler, 1956; Patterson, 1956; Vanderbroek, 1961; Simpson,1961a; Weller, 1968).
К этому времени стало известно, что примитивнейшие цинодонты как раз и обладали заклыковьгми зубами упрощенного гриконодонтного типа. Это возродило оптимизм исследователей, пытающихся доказать монофилетическое происхождение млекопитающих (Parrington, 1967b; Hopson, Crompton, 19 69), и стимулировали широкое переизучение материалов по зубам их мезозойских представителей ( Crompton, Hilemae,1960; Crompton, Jenkins, 1967; 1968;Jenkins, .1969a; Mills, 1971; Parrington, 1971, 1973; Kennack et.al., 1968; Kermack ее аl, 1973). Эти работы, проводившиеся преимущественно на многочисленных, но изолированных зубах, стали возможными после тото, как Майлс (Mills,. 1964), Эвери и Кюне Every, Kuhne, 1971) показали возможность 'гомологии" фасеток истирания на бугорках коренных зубов млекопитающих. Каждая фасетка на протокониде у териевых млекопитающих, например, связана с изнашиванием при прикусе с соответствующей поверхностью паракона или метакона. Каждый верхнекоренной зуб верхнетриасовых млекопитающих: имеет не менее восьми фасеток (Crompton, Jenkins, 1968). В результате удалось создать довольно стройную концепцию эволюции коренных зубов основных групп млекопитающих и показать, что зубы симметродонтов и пантотериев легко выводятся из зубов верхнетриасовых 'триконодонтов' типа Kuhneotherium (Kermack etal.,1968;Crompton, 1971).
Все эти группы вместе с сумчатыми и плацентарными объединяются теперь обычно в подкласс Theria (Kermack,1967b; Hopson, Crompton, 1969). С другой стороны, зубы типичных юрских триконодонтов и докодонтов легко выводятся из зубов верхнетриасового Morganucodon (Kermack, 1967), однако фасетки истирания на зубах у кюнеотериид и морганукодонтид оказались негомологичными. Прикус в обеих этих группах мезозойских млекопитающих устанавливался параллельно. Функционально зубная система у всех верхнетриасовых млекопитающих находится на режущей стадии эволюции прикуса ( Butler, 1972), но у кюнеотериид протоконид при смыкании челюстей занимает переднее положение - между параконом и метаконом верхних моляров, а не позади метакона, как у морганукодонтид (Mills, 1971; Crompton,1971; Parrington, 1971). Кюнеотериид на основе этих данных выделили из состава триконодонтов; Кромптон и Дженкинс относят их непосредственно к симметродонтам (Crompton, Jenkins, 1973).
Результаты исследования зубов мезозойских млекопитающих не дали решающих аргументов в пользу их полифилетического или монофилетического происхождения. Общее морфологическое сходство зубов морганукодонтид и кюнеотериид свидетельствует все же в пользу относительно недавней дивергенции обеих групп от цинодонтовых предков. Паррингтон (Parrington, 1967b, 1971), Хопсон, Кромптон и Дженкинс (Hopson, Crompton, 1968; Hopson,1969, 1970} Crompton, Jenkins, 1973) склоняются к выводу о монофилии млекопитающих. Паррингтон, например, склонен видеть общих предков млекопитающих в формах, близких к диартрогнатам. От верхнетриасовых 'триконодонтов', по Паррингтону, происходят не только симметродонты и пантотерии, но и многобугорчатые млекопитающие. Связующее звено между обеими группами Паррингтон ( Parrington, 1967 b) видит в харамийидах. В то же время Паррингтон солидаризуется с Хопсоном ( Hopson,1966) во мнении о параллельном приобретении "маммального" звукопроводящего аппарата клоачными млекопитающими и териями, в пользу чего говорит хотя бы наличие у них особого депрессора нижней челюсти (m. decrahens mandibulae), не гомологичного ни рептилийному m. depressor mandibulae, нитериевому т. digastricus. Замена рептилийного депрессора происходила у предков обеих групп параллельно в связи с преобразованием двойного челюстного сустава предков млекопитающих в "маммальный".
Иных взглядов придерживается Кермак, защищающий гипотезу дифилетичес-кого происхождения млекопитающих (Kermack,19б7а,Ь, 1971). Кермак акцентирует внимание на том, что у всех известных в этом отношении "нетериевых" групп предушная стенка эндокрания образована не алисфеноидом, как у сумчатых и плацентарных, а выростом периотики, как у монотремат (Morganucodon: Kermack et al.,1957; Sinoconodon: Pacterson, Olson, 1961; типичные юрские триконодонты: Kermack, 1963б мультитуберкуляты: Kielan—Jaworowska, 1971); только у типичных юрских докодонтов эта особенность эндокрания осталась неисследованной. Ганглий тройничного нерва у всех этих форм расположен на внутренней поверхности переднего отростка периотики, образующего, по мнению названных авторов, наружную стенку эпиптериковой полости у всех нетериевых млекопитающих. На этом основании делается вывод, что териевые и "нетериевые" млекопитающие происходят самостоятельными стволами, дивергировавшими еще на уровне архаичных цинодонтов, не приобретших еще предушной пластины периотики (Kermack, Kielan—Jaworowska, 1971; Kermack et al.,1973).
Мнение о возможности противопоставления 'нетериевых' млекопитающих териевым получило широкое признание. Соответственно "нетериевых" млекопитающих объединяют в подклассе Prototheria (Hopson, 1970; Crompton, Jenkins, 1973), Atheria (Kermack, et al,, 1973) или даже выделяют из состава млекопитающих под названием 'квазимлекопитающих" (Mac Incyre,1967; см. также: Fox, 1967). Однако если генетическое единство подкласса Theria можно считать хорошо обоснованным, то этого нельзя, по нашему мнению, сказать о 'нетериевых" млекопитающих. Передний отросток периотики был хорошо развит даже у примитивных териодонтов, и у цинодонтовых предков млекопитающих стенка черепа в предушной области была образована как этим выростом, так и восходящей пластиной эпиптеригоида. При этом вырост периотики пересекал эпиптериковую полость, подразделяя ее на дорсомедиальную и вентро-латеральную части, а не выстилал ее изнутри; ганглий тройничного нерва и у териодонтов располагался, по-видимому, на внутренней стенке переднего отростка периотики (см. рис. 18). Превращение двустенной предушной эндокра-ниальной стенки в одностенную проходило в результате уменьшения размеров эпиптериковой полости, связанного с увеличением размеров головного мозга и оттеснением наружу внутренней стенки эпиптериковой полости. В результате, вырост периотики, образующий "интраэпиптериковую" перегородку, постепенно сближался с восходящей пластиной эпиптеригоида, превращавшейся у териевых форм в алисфеноид. У известных в этом отношении прототериев вырост пе— риотики заместил восходящую пластину апиптеригоида, причем не исключено, что это замещение проходило параллельно в различных линиях прототериев. По строению зубов родство с верхнетриасовыми млекопитающими мультитуберку-лят и монотремат нельзя считать достаточно обоснованным; не исключено поэтому, что обе эти группы примитивных млекопитающих могли развиваться от цинодонтового уровня самостоятельно. Теоретически нельзя исключать и возможность родства их с архаичнейшими териями типа кюнеотериид, которые в таком случае должны были наряду с алисфеноидом обладать и хорошо развитой передней пластиной периотики и быть в этом отношении более примитивными, чем морганукодонтиды. В связи с этим небезынтересно напомнить, что Кюне ( Kiihne, 1973) и сейчас предпринимает попытку доказать тесное родство монотремат с сумчатыми млекопитающими, предполагавшееся Грегори (Gregory, 1947), а Ромер (Romer,1970) скептически относится к попыткам связать монотремат, мультитуберкулят и харамийид с какой бы то ни было группой верхнетриасовых млекопитающих, хотя высказывается в пользу монофилии этого класса.
Вряд ли какие бы то ни было млекопитающие могут происходить от диартрогнатов. Кромптон и Дженкинс ( Crompton, Jenkins,1968) указывают, что архаичнейшие из млекопитающих - триконодонты - по строению зубов могут выводиться лишь из наиболее примитивных цинодонтов типа Thrinaxodon. Диартрогнаты по строению зубов явно стоят в стороне от предков основной (первой) группы млекопитающих. По нашему мнению, однако, еще ближе к верхнетриасовым млекопитающим по строению зубов стоят более примитивные цинодонты - проиинозу-хиды или менее специализированные галезавриды типа Nanocynodon. У этих форм зубы менее расширены поперечно, чем у тринаксодона, главная вершина их сильнее выдается над боковыми, а число придаточных бугорков на линг-вальном пояске не так велико и не превышает четырех-пяти. Из данных Кромп-тона и Дженкинса явствует, что особым сходством с триконодонтами обладают зубы лишь ювениальных особей тринаксодона (см. рис. 21), у взрослых особей которого задние зубы построены явно сложнее.
Вывод о родстве верхнетриасовых млекопитающих с галезавроидеями исключает наиболее 'маммальных" териодонтов из числа предков основной группы млекопитающих. Становится очевидным, что эволюция в направлении млекопитающих совершалась у высших гомфодонтных цинодонтов (включая тритило— донтов) и диартрогнатов совершенно независимо от реальных предков млекопитающих. Из числа высших цинодонтов не исключено родство с млекопитающими лишь рода Probainognathus, выделяемого Ромером в Probainognathidae (Romer, 1970, 1973,», а также некоторых форм, объединяемых нами в Dromatheriidae. Пробайногнат сближается с предками млекопитающих наличием дополнительного нижнечелюстного сочленения между зубной и чешуйчатой костью, однако заклыковые зубы, характеризующиеся очень слабо выраженной главной вершиной и наличием лишь зачаточного пояска (Romer, 1970, рис. 9;, на наш взгляд, отделяет эту форму от прямых предков верхнетриасовых млекопитающих. Высказывания Ромера (Romer, 1970, 1973), о близости пробайногната к предкам млекопитающих, видимо, недостаточно обоснованы. Против родства пробайногната с млекопитающими говорит и практически полная редукция у него ангулярного лепестка, остававшегося палочковидным у ряда других высших цинодонтов и сравнимого, по-видимому, с нижней развилкой тимпанальной кости млекопитающих, поддерживающую барабанную перепонку. Ромер склонен считать, что нижняя развилка тимпанальной кости появилась у млекопитающих вторично. Против этого, однако, свидетельствует крайнее ее сходство с преобразованной ангулярной пластиной диартрогнатов (см. рис. 25} и некоторых других цинодонтов. Поэтому мы склонны считать высших цинодонтов с крайне выраженной редукцией ангулярного лепестка стоящими в стороне от прямых предков млекопитающих.
То обстоятельство, что высшие териодонты приобрели характерные для млекопитающих особенности, не должно затенять сохранения в их организации многих архаических признаков, уникальных для типичных пресмыкающихся. Как маммальные, так и архаические особенности в различной степени выражены у разных териодонтов. Широкий размах процесс маммализации приобретает, пожалуй, только у высших териодонтов, прежде всего у цинодонтов и в меньшей степени у бауриоморфов.
Локомоция.
Только низшие звероообразные - пеликозавры - сохраняли в полной мере обычное для примитивных тетрапод положение проксимальных отделов конечностей в горизонтальной плоскости. Во всех ветвях терапсид происходит хотя бы в зачаточной степени поворот конечностей под тело - в парасагиттальную плоскость, что значительно повышает эффективность их функционирования. Выраженной степени этот поворот достигает у высших териодонтов - у бауриоморфов и цинодонтов. У примитивных цинодонтов типа Thri— пахоаоп плечо направлено под углом 45°, а бедро - под углом 55° к продольной оси тела по сравнению с 25—35° у примитивных млекопитающих и 80° у пеликозавров (Jenkins, 1970 a,b,c;l971).
Имеются основания говорить о постепенном приобретении прогрессивными терапсидами, особенно териодонтами, и многих других характерных особенностей локомоторного аппарата млекопитающих. У большинства териодонтов исчезает клейтрум, лопатка приобретает ость, хотя ее супраспинаторная яма не развита даже у прогрессивных цинодонтов. Гленоидная впадина постепенно смещается назад и у типичных бауриоморфов, цинодонтов и даже дицинодонтов обращена не наружу, как у пеликозавров, а постеровентрально. У всех этих терапсид гленоидная впадина целиком ограничена лопаткой и метакораксидом, передний коракоид у высших цинодонтов (траверзодонтиды типа Exaeretodon и тритилодонты) сильно редуцирован, форма гленоидной впадины из винтообразной, свойственной пеликозаврам и всем низшим тетраподам, становится округлой у всех высших терапсид, что дает плечу значительно больший простор движений и свидетельствует об усложнении нейромышечного контроля над поддержанием тела и локомоцией. По строению плечевого пояса к цинодонтам очень близки монотрематы, также сохраняющие небольшой передний коракоид и не и-меющие супраспинаторной ямы на лопатке (Watson, 1917; Gregory, Camp, 1918; Romer,1922; Boonstra, 1968Ь; Jenkins,, 1970а ).
Еще более приближаются высшие териодонты к млекопитающим по строению тазового пояса, эволюирующего в маммальном направлении быстрее плечевого (Crompton, Jenkins, 1973). Отметим здесь характерное особенно для цинодонтов переднее разрастание подвздошной кости, принимающей антеродорсальную ориентацию, увеличение числа крестцовых позвонков до пяти, происходящую у высших цинодонтов (Exaeretodon) и тритилодонтов редукцию заднего отростка подвздошной кости, укорочение лонной кости и резкое расширение обтураторного отверстия, превращающегося в широкое окно маммального типа. По крайней мере у тритилодонтов имелись сумчатые, или предлобковые, кости. Верт-лужная впадина постепенно переходит из латерального положения в нижнее (Gregory, Camp, 1918;Romer, 1922; Boonstra, 1968 b; Jenkins, 1970 а).
Проксимальные кости конечностей у терапсид, особенно у териодонтов, постепенно становились более длинными и тонкими, а Проксимальные сочленовные поверхности плечевой и бедренной костей округлыми. Однако эпифизы длинных костей не отделялись у териодонтов хрящевыми зонами от диафизов.
Плечевая кость оставалась относительно массивной, но дельто-пекторальный гребень у цинодонтов и бауриоморфов несколько уменьшался в размерах. Сохранялись еще оба дистальных надмыщелка и оба эпикондилярных отверстия; эти признаки свойственны и большинству архаичных млекопитающих, в частности монотрематам. Локтевая кость приобретала длинный проксимальный отросток - олекранон. Лучевая кость, по крайней мере у цинодонтов, была пронирована относительно локтевой, так что дистальные концы обеих костей перекрещивались. Число карпальных костей сокращалось до 11 у цинодонтов (Thrinaxodon: Romer, 1922; Boonstra, 1968 b; Jenkins, 1970a).
В бедренной кости у цинодонтов намечалось обособление проксимальной головки от тела кости. Примитивная Y-образная система аддукторных гребней постепенно перестраивалась, преобразуясь в вертлуги (трохантеры); у высших териодонтов появлялся большой вертлуг маммального типа, а внутренний редуцировался, возможно преобразуясь в малый вертлуг, который появляется только у цинодонтов. Такое изменение внутреннего вертлуга было сопряжено с некоторой перестройкой в составе прикреплявшихся к нему мышц. У высших териодонтов астрагал и пяточная кости преобразуются по маммальному образцу, причем появляется хорошо выраженный пяточный бугор. Общее число костей стопы уменьшалось до шести—семи, причем стопа становилась сводчатой. Эти изменения особенно резко были выражены у бауриоморфов, опережавших в этом отношении цинодонтов (Romer, 1922; Parrington, 1961; Boonstra, 1968b; Schaeffer, 1941; Jenkins, 1970a).
Поворот проксимальных отделов конечностей под тело был сопряжен с упрощением фаланговой формулы, которая приближалась к маммальной у дицинодонтов, высших тероцефалов, бауриоморфов и высших цинодонтов (Boonstra, 1968 Ь). Исчезновению 'избыточных* фаланг III и IV пальцев предшествует их резкое укорочение - даже у горгонопсов вторая фаланга III пальца и вторая и третья фаланги IV пальца резко укорочены (Colbert, 1948). Сокращение числа фаланг до маммального совершалось в различных группах терапсид параллельно; даже у примитивных цинодонтов (Thrinaxodon) фаланговая формула была еще почти рептилийной - 2, 3, 4, 4. 3, тогда как у прогрессивных она становилась маммальной (Jenkins, 1970 а). Изменялась и длина пальцев - самым длинным постепенно становился не четвертый, а третий; иногда все пальцы, кроме первого, выравнивались по длине (тероцефалы, дейноцефалы: Boonstra, 1968 b).
Не исключено, что перестройка локомоторного аппарата сопровождалась выработкой нового типа координации движений с переходом от примитивной симметричной покомоции к асимметричной, характерной для млекопитающих. Эти изменения были важнейшим событием в эволюции локомоторного аппарата наземных позвоночных (Суханов, 1968). Косвенное указание на приближение по этому признаку к млекопитающим дает характерное для всех терапсид и особенно для териодонтов значительное увеличение размеров мозжечка, что выражается в появлении и прогрессирующем увеличении размеров флоккулярной ямы на внутренней стенке периотики. Все это позволяет говорить о значительном повышении активности зверообразных, что рассматривается иногда в качестве наиболее характерной особенности эволюции териодонтов (Olson,1959; Brink, 1963a). Дополнительным подтверждением маммального характера некоторых по крайней мере особенностей локомопии высших териодонтов дает также описанная Бринком ( Brink, 1959a) находка цинодонта Thrinaxodon Нот— hinus, захороненного в типичной для млекопитающих позе - свернувшегося клубком. Однако в полной мере маммальный механизм координации движений, морфологически выражающийся в развитии двигательной зоны коры больших полушарий головного мозга, связанного с ней так .называемого пирамидного тракта и полушарий мозжечка, видимо, был приобретен лишь на более поздних этапах эволюционного развития. Во всяком случае большие полушария у териодонтов -оставались очень маленькими, а пирамидный тракт и полушария мозжечка находятся на крайне примитивной ступени развития даже у современных клоачных млекопитающих (Bautista, 1968).
О том, что маммальная "симметричная" локомоция, осуществляемая при участии вертикальных изгибов тела, еще не была до конца выработана териодонтами, свидетельствует отсутствие у них выраженной дифференциации туловищных позвонков в отношении длины остистых отростков, столь характерной для млекопитающих. Даже у прогрессивных цинодонтов такая дифференциация только намечается. Туловищный отдел позвоночника у териодонтов не имел двух характерных для млекопитающих изгибов - шейного и спинного и в покое позвоночник был прямым (Jenkins, 1971b). Наконец, расширенность проксимальных отделов ребер цинодонтов говорит о мощном развитии у них системы подвздош-но—реберных мышц, играющих важную роль при боковых изгибах тела у рептилий. У циногнатий эти реберные пластинки могут даже сочленяться друг с другом. Это обстоятельство, однако, дает основания для предположения, что реберные пластинки стабилизировали положение туловищного отдела позвоночника, подобно тому как это имеет место у некоторых современных роющих и древесных неполнозубых, также обладающих расширенными реберными пластинками, не связанными, однако, с системой подвздошно—реберных мышц (Jenkins, 1970b).
Питание.
Дифференциация зубов у скалопоэавров и цинодонтов определенно указывает на переход их от заглатывания пищи большими кусками к ее измельчению или даже пережевыванию в ротовой полости. Характерный для низших териодонтов способ питания был, по-видимому, единственно возможным для форм с редуцированными заклыковыми зубами (иностранцевииды, рубиджевиды среди горгонопсов и вайтсииды и эвчамберсииды среди тероцефалов;. Это подтверждается отчасти и находкой экземпляра вайтсиида Notosollasia boonstrai с черепом дицинодонта между ветвями нижней челюсти; очевидно, хищник погиб, подавившись слишком большим куском (Brink, 1959)
Среди мелких хищных териодонтов с трехзубчатыми заклыковыми зубами (иктидозухи, галезавроидеи и некоторые другие) имелись, по-видимому, и формы, питавшиеся насекомыми. Насекомоядными в значительной мере были, вероятно, и предковые для млекопитающих териодонты (Родендорф, 1972).
Значительно облегчало обработку пищи в ротовой полости приобретение высшими териодонтами вторичного костного нёба. Первично это нёбо укрепляло челюстной аппарат. В ряде случаев оно использовалось в качестве платформы для дробления на нем пиши нижними заклыковыми зубами. Эта особенность характерна для хищных цинодонтов, у которых коронки заклыковых зубов при смыкании челюстей не соприкасались друг с другом. В наибольшей степени такое дробление пищи, возможно содержащей крупных насекомых с твердым хитиновым панцирем, было свойственно северодвинским цинодонтам (Татаринов. 1974).
Среди высших териодонтов с полным вторичным костным нёбом наблюдается значительная радиация форм, перешедших к растительному питанию (баурииды, гомфогнатоидеи, тритилодонты, диартрогнаты).
Среди гомфогнатоидеи наблюдаются необычайное усложнение и многообразие типов распределения придаточных бугорков на жевательных поверхностях заклыковых зубов. У них и бауриид впервые в эволюции териодонтов устанавливается смыкание (окклюзия) заклыковых зубов своими жевательными коронками, резко повышающими эффективность их работы при перетирании пищи. Поскольку окклюзия зубов устанавливается у териодонтов на базе весьма усложненных жевательных коронок, эволюировавших до того независимо друг от друга, на ранних этапах эволюции в функционировании зубной системы у таких форм имелось немало дисгармоничных черт. В ряде случаев у примитивных траверсодонтид например, бугорки верхних зубов смыкались не с ложбинками, а бугорками нижних; такие зубы подвергались очень быстрому изнашиванию. В эволюции среднетриасовых траверсодонтид удалось проследить постепенное совершенствование окклюзии зубов» становившейся в итоге вполне гармоничной ( Crompton,1972).
Наибольшего усложнения коронки зубов достигали у тритилодонтов, у которых бугорки на верхних заклыковых зубах формировали три, а на нижних - два продольных ряда. При пережевывании пиши преобладали переднезадние движения нижней челюсти. Тритилодонты в известной мере предваряют экологический тип грызуна, что подтверждается, в частности, долотовидностью их резцов и утратой клыков. Они были непосредственными экологическими предшественниками многобугорчатых млекопитающих и вымерли в средней юре, по-видимому в условиях конкуренции с предками мультитуберкулят.
Косвенное указание на приспособление к обработке пищи в ротовой полости дают признаки развития у териодонтов серии щечных желез (предглазничная, назо-челюстная, челюстная), возможно сравнимых со слюнными. По крайней мере часть этих желез была приобретена еще низшими териодонтами (горгонопсами и тероцефалами).
Дыхание.
Формирование у эвтериодонтов (включая тероцефалов) выраженного поясничного отдела позвоночника говорит о приобретении ими диафрагмы и перехода к маммальному механизму вентиляции легких (Attridge, 1956; Brink, 1957а) . Добавим, что происхождение диафрагмальных мышц млекопитающих главным образом за счет шейных и ее иннервация шейным нервом (С 6) говорит о появлении хотя бы зачаточной диафрагмы еще у предков зверообразных с практически неразвитой шеей. Возможно, что диафрагмаль-ное дыхание эффективнее рептилийного "реберного"'. Тогда формирование диафрагмы и обособление поясничного отдела позвоночника косвенно указывает на прогрессирующее повышение активности териодонтов.
Головной мозг и органы чувств. Примитивные териодонты (горгонопсы) отличаются совершенно неразвитым передним мозгом. В этом отношении они резко отстают от любых современных пресмыкающихся. Самым развитым отделом головного мозга у горгонопсов был, по-видимому, мозжечок.
Значительное увеличение размеров переднего мозга отмечается только у высших териодонтов. Скудные сведения о размерах у них черепной полости не позволяют вынести окончательного суждения, но создается впечатление, что у тероцефалов и бауриаморфов (Татарине в, 1974) и типичных цинодонтов ( Watson, 1913Ь; Bonaparte, 1966 ) передний мозг оставался очень маленьким по сравнению с современными пресмыкающимися и даже мезозойскими млекопитающими — триконодонтами (Siinpson.1927).
Своеобразие строения головного мозга зверообразных подчеркивается сохранением у большинства их глазоподобного пинеального органа, как у бесхвостых земноводных и круглоротых, а не париетального, как у современных пресмыкающихся.
Из органов чувств отмечается необычайное развитие обонятельного. Носовые капсулы занимают у зверообразных от трети до половины общей длины черепа. По крайней мере у териодонтов имеются четкие признаки развития верхних обонятельных раковин (назотурбиналий). Якобсонов орган, как и у современных млекопитающих, открывался, по-видимому, в ротовую полость через остатки переднего нёбного окна.
Нет данных, которые позволяли бы определенно говорить о прогрессе органа слуха. Характерное для млекопитающих выпячивание под кохлеарным отделом внутреннего уха — промонторий — не выражено у териодонтов. По нашему мнению, настоящая барабанная перепонка у типичных зверообразных отсутствовала, и слух был у них пониженным. В качестве зачаточной барабанной перепонки у большинства зверообразных могла использоваться утоньшенная стенка дна ротовой полости в области мандибулярного дивертикула барабанной полости, медиальнее задней части нижней челюсти. Колебания дна ротовой полости могли передаваться слуховой косточке через посредство рожков подъязычной кости. Только у цинодонтов, по-видимому, начинала формироваться настоящая барабанная перепонка, прикреплявшаяся к преобразованному ангулярному лепестку нижней челюсти и, возможно, погруженная глубоко под кожу и мышцы. С внешней средой она сообщалась посредством длинного и изогнутого хрящевого наружного слухового прохода, подобно тому как это имеет место у монотремат. Возможно, что повышение остроты слуха, сопряженное с развитием барабанной перепонки, нашло свое отражение в преобразовании зрительных долей в характерное для млекопитающих четверохолмие.
Кожа, водный обмен, сердце. Особенности водного обмена млекопитающих, в частности сохранение у них гломерулярной почки и выделение в качестве основного конечного продукта белкового обмена не мочевины, а мочевой кислоты, говорят об их происхождении от форм, обладавших амфибийным типом водного обмена (Smith, 1953). Это наводит на мысль, что богатство кожи млекопитающих железами и отсутствие развитого чешуйчатого покрова унаследованы ими в конечном счете еще от земноводных (Татаринов, 1959).
Полученные сведения о строении кожи у зверообразных в известной мере подтверждают такое предположение. Чудиновым {1970) описаны отпечатки кожи дейноцефала Estemmenosuchus, лишенные всяких признаков роговых чешуи рептилийного типа, но содержащие многочисленные чечевицеобразные структуры, по-видимому, железистой природы. Вся эпидермальная часть кожи очень тонка.
Не исключено, что тонкая и богатая железами кожа, по крайней мере у •наиболее примитивных зверообразных, еще была проницаемой и играла некоторую роль в дыхании.
Особенности строения сердца у современных млекопитающих говорят о самостоятельном переходе их от трехкамерного амфибийного сердца к четырехкамерному, что, возможно, связано с переходом от кожно-легочного 'амфибийного' дыхания к легочному (Татаринов, 1960).
Эти наблюдения показывают, что по крайней мере примитивные зверообразные были относительно слабо защищены от иссушающего действия воздуха. Такая защита достигалась у них в значительной мере не толстым орого-вевающим эпидермальным покровом, как и у современных пресмыкающихся, а его увлажнением сильно развитыми кожными железами. Дополнительное указание на это дает развитие у зверообразных массивной латеральной носовой железы, выполняющей функцию увлажнения носовой полости. В этом отношении зверообразные сохраняли еще некоторое сходство с амфибиями.
Губы, щеки, лицевая мускулатура и вибриссы.
Как уже отмечалось, имеются косвенные признаки развития в губной области высших териодонтов сенсорных придатков типа "протовибрисс", а у цинодонтов (и, возможно, баурио-морфов; также и мягких 'мускульных* губ маммального типа (Татаринов, 1967). Напомним здесь обнаруженные нами у Dvinia prima вероятные отпечатки губных мышц, резорбцию межносового костного мостика у Diademodon (Brink, 1963d) и, наконец, полное исчезновение этого мостика у тритилодонтов. Последовательные ступени развития этих признаков наблюдаются у различных высших териодонтов.
Проблема теплокровности.
Признаки прогрессирующего повышения активности в эволюции териодонтов, развития вибрисс, вторичного неба, зубов, способных к пережевыванию пищи и, наконец, утрата пинеального отверстия привели некоторых исследователей к мнению, что высшие териодонты в той или иной мере становились теплокровными ( Brink, 1963а)» Наиболее категорически это высказано Вьюшковым (1951, 1956) и Ефремовым (1954, 1961), которые считают признаками настоящей теплокровности "высших териодонтов развитие у них вторичного костного нёба и редукцию теменного глаза - "органа фотогормональной терморегуляции'. О теплокровности поздних териодонтов говорит, по Вьюшкову, и появление в конце перми ядовитого тероцефала Euchambersia; по его мнению, ядовитые животные биологически эффективны лишь при наличии в фауне большого числа теплокровных форм, ибо на холоднокровных животных яды обычно действуют медленнее, чем на теплокровных.
На наш взгляд, все эти аргументы совершенно недостаточны для суждения о температурном режиме териодонтов. Мы уже отмечали, что развитие вибрисс само по себе еще не означает наличия развитого волосяного покрова, способного к выполнению термоизолирующей функции (Татаринов, 1965а). Возможно, что эволюционно волосяной покров млекопитающих развивался на основе именно сенсорных волосков (По мнению Коупса (Cowles,1946, 1957), первичной функцией волосяного покрова у предков млекопитающих была защита от перегрева). Редукция теменного глаза, видимо, вообще не связана с теплокровностью, а ядовитыми становятся и многие беспозвоночные, рыбы и некоторые змеи семейства Colubridae, не питающиеся теплокровными позвоночными. Развитие аппарата пережевывания пищи действительно связано с повышением активности, но не дает оснований для заключеяия о теплокровности. Еще менее доказательно в этом отношении развитие вторичного костного неба, первые этапы развития которого у териодонтов связаны не с изоляцией пищеварительного тракта от дыхательного, что не крайне важно для теплокровных животных, а с механическим укреплением челюстного аппарата, связанного с переходом к пережевыванию пищи.
Маловероятно, что даже высшие териодонты были настоящими теплокровными. Кромптон (Crompton, 1968а) считает, что и у всех мезозойских млекопитающих терморегуляция была еще несовершенной: по его мнению, эти животные могли поддерживать температуру тела на более высоком уровне, чем температура среды, но не были способны эффективно повышать теплоотдачу при необходимости защиты от перегрева. Даже у современных клоачных и некоторых сумчатых млекопитающих температура тела колеблется в широких пределах, порядка 10°. Особенно интересны в отношении терморегуляции однопроходные, среди которых у ехидны нормальная температура тела составляет всего 30-31° (на 7-8° меньше, чем у плацентарных). При значительном повышении температуры воздуха ехидна не способна поддерживать постоянную температуру тела и "перегревается" (Schmidt—Nielsen ее al.,1966).
Против теплокровности териодонтов косвенно говорит и отсутствие у них каких-либо признаков развития нижней (челюстной обонятельной раковины, . тогда как гребни для верхней хорошо представлены, по-видимому, у всех териодонтов. Напомню, что у млекопитающих нижняя раковина в отличие от верхней покрыта не обонятельным, а дыхательным эпителием, выполняя функцию обогрева и увлажнения вдыхаемого воздуха, и обычно развита намного сильнее верхней. Таким образом, нижняя обонятельная раковина связана с развитием теплокровности, а верхняя — с интенсификацией обоняния.
Утверждение Ван Валена (Van Valen, 1960), что у териодонтов обычно имеются признаки развития нижней обонятельной раковины, поддерживаемой септо-максиллой и мезэтмоидом, ошибочно. Основанием для него послужило высказанное в прошлом предположение Уотсона (Watson, 1913b) о связи интермедиального отростка септомаксиллы пинодонтов с нижней челюстной раковиной. Теперь, после исследований септомаксиллы низших позвоночных Ярвиком (Jarvik, 1942), такое высказывание полностью утратило смысл.
Рикле (Riqlet, 1969, 1972) пришел к выводу о теплокровности терапсид (и динозавров) на основании некоторых особенностей гистологии их костной ткани: отсутствия годичных колец, значительного сходства гаверсовых систем и наличия первичной пластинчатой костной ткани. Первый признак указывает, однако, лишь на возможную круглогодичную активность исследованных Рикле животных. Значение остальных признаков остается неясным.
О размножении. Единичные данные по этому вопросу не позволяют прийти к определенному выводу. Отметим все же, что высказывание Конжуковои (1949} о возможной, судя по большому размеру тазовой полости, живородности северодвинских цинодонтов основано на недоразумении. При прочих равных условиях откладывание яиц, в котором зародыш свернут клубком, требует больших размеров тазовой полости, чем живородность.
Бринк ( Brink, 1965а) описывает очень интерсную находку "семьи"' Thrinaxodon, состоящую из черепа взрослой особи, по-видимому самки, и черепа очень маленькой ювенильной особи. Оба образца заключены в одну конкрецию. Нельзя считать доказанной данную Бринком интерепретацию этой находки в качестве семьи, но возможно, что эта находка действительно указывает на развитие высокой формы заботы о потомстве у цинодонтов.
Бринк (Brink, 1957) отмечает также, что "новорожденные* особи Diademodon, возможно, обладали лишь немногочисленными и коническими заклыковыми зубами. Это явно делает их не способными к питанию пищей взрослых животных и, видимо, указывает на известное приспособление к вскармливанию молоди молоком. Однако эта идея сильно дискредетирована тем фактом, что у наиболее молодых особей диадемодона немногочисленные заклыковые зубы были, по-видимому, сложно дифференцированы. Они замещались просто устроенными субконическими вторично, по мере удлинения зубного ряда (Hopson, 1971). Во всяком случае независимо от устройства зубной системы у 'новорожденных" сосание молока стало возможным с момента приобретения предками млекопитающих губной мускулатуры.
Необходимо отметить, что открытие у тритилодонтов сумчатых костей (Fourie, 1963) сделало вероятным предположение о наличии у них и сумки, в которой эти териодонты могли вынашивать яйца, подобно тому как это делает ныне ехидна. Возможно, что млечные железы, первоначально развившиеся, по-видимому, в паховой области и генетически связанные с потовыми, первоначально имели функцию не вскармливания молоди, а создания оптимального микроклимата в сумке посредством увлажнения ее содержимого (Hopson,1973^ В таком случае переход к молочному питанию мог совершаться очень постепенно, через стадию, на которой вылупившиеся из яйца "новорожденные" особи слизывали жидкие выделения "протомолочных" желез, постепенно обогащавшихся питательными веществами. Может быть, первоначально эти железы служили в большей мере источником воды и солей, а не пищи в прямом смысле слова.
Заключение.
В организации зверообразных прогрессивные "маммальные" особенности сочетаются с крайне примитивными. Из числа таких примитивных особенностей отметим: подразделение мозговой коробки на два отдела;слабое развитие больших полушарий головного мозга; железистая и, возможно, проницаемая кожа; строение слезного протока с очевидным включением в него части инфраорбитального канала боковой линии; сохранение переднего нёбного овна; дифференциация сосудов и нервов в предглазничном отделе черепа.
Некоторые особенности современных млекопитающих дают основание говорить об обособлении аверообрааных от предков, еще сохранявших амфибийный тип водного обмена и амфибийное средне с неподразделенным желудочком. При этом если в организации высших териодонтов на первый план выходят "маммальные" особенности, то в организации наиболее примитивных пеликозавров превалируют архаические признаки. Эти обстоятельства делают малоуспешными попытки дать единую морфофизиологическую характеристику зверообразным пресмыкающимся, аналогичную существующим для земноводных современных пресмыкающихся, птиц и млекопитающих. Вряд ли достаточна здесь формула Бринка "преодолевающие термальный берьер" (Brink, 1963 а), поскольку она приложила, и то не в полной мере, лишь для высших 'эвгериодонтов". По этой же причине неосновательны и предположения, по которым все вообще териодонты находились на "протериевом" физиологическом уровне (Carter, 1957). Совершенно бесплодны также попытки приспособить к зверообразным морфофизиологическую характеристику современных пресмыкающихся. Следование такому ложноактуалистическому методу приводило к сравнению зверообразных с крупными ящерицами. Быстров (1955; прямо называл териодонтов 'хищными ящерицами', и это представление по существу испортило технически безукоризненно выполненные им реконструкции внешнего облика зверообразных пресмыкающихся, моделью для которых послужили современные ящерицы (Быстров, 1933).
Низшие зверообразные, появляющиеся уже в конце среднего карбона (Protoclepsidrops: Саrгоll,1964), смыкаются с архаичнейшими наземными позвоночными. В целом подкласс Theromorpha (=Synapsida) правильнее рассматривать как заполняющий огромный пробел между ранними тетраподами и млекопитающими, но не как связующий типичных завропсидных пресмыкающихся с млекопитающими. В этом смысле можно говорить о возрождении в наши дни прежних идей Т.Гексли (Huxley, 1880) о происхождении млекопитающих от земноводных.