
Проблемы Эволюции |
О взрывном видообразовании, протекающем на наших глазах в высыхающем Аральском море. Омск: Изд-во Омского государственного педагогического университета, 2003. 382 с.
Глава 2. КРАТКАЯ ИСТОРИЯ ФОРМИРОВАНИЯ АРАЛЬСКОГО МОРЯ И ПУТИ ПРОНИКНОВЕНИЯ ДВУСТВОРЧАТЫХ МОЛЛЮСКОВ
Изложение истории формирования Аральского моря и его фауны следует предварить словами Д.Д. Квасова (1991): «Сложная история Арала до сих пор не понята до конца» (с. 219). Несмотря на огромное количество статей и книг, посвященных этой проблеме, воззрения на историю формирования Аральского моря и его фауны до сих пор противоречивы и туманны. Различны датировки возникновения и самого водоема и проникновения в Арал отдельных элементов его фауны. Одно несомненно, что основной чертой геологической истории Приаралья является частая смена водоемов с водой различной солености и разного ионного состава.
По материалам последних исследований история современного Аральского моря невелика – оно возникло на рубеже плейстоцена и голоцена, в результате заполнения Арало-Сарыкамышской котловины водами Амударьи и Сырдарьи, достигшими Арала всего лишь около 10 тыс. лет назад (Квасов, 1991). Вся история существования Аральского моря представляет собой серию неоднократно сменявших друг друга трансгрессивных и регрессивных стадий, во время которых наступавшее или отступавшее море имело разную соленость и соответствующую ей биоту. Так, на основании изотопно-кислородного анализа было установлено, что до голоцена на месте центральной котловины существовали небольшие пересыхающие водоемы с резко колеблющейся соленостью. Эта озерно-солончаковая стадия развития Арала сменилась в голоцене стадией, характеризуемой наличием зеленовато-серых морских осадков, характерных для солоноватых водоемов, включающих раковины Cerastoderma (Маев с соавт., 1983а, б; Квасов, 1991; Маев, Маева, 1991; Николаев, 1991; и др.).
Довольно значительное повышение уровня по изотопно-кислородным данным происходило между 4.8–3.6 тыс. лет, максимальная регрессия имела место 1600 лет назад, а последняя регрессивная фаза зарегистрирована примерно 600 лет назад. В период максимальной регрессивной стадии в центральной котловине существовал проточный водоем с неоднородной соленостью, сменяющейся по акватории почти от нулевой до нескольких промилле, позволявшей существовать кардиидам наряду с доминировавшей пресноводной фауной (Маев, 1983; Николаев, 1991).
До сих пор остается неясным вопрос датировки наиболее древних аральских террас 73 и 53–58 м над уровнем океана. Наиболее обширная трансгрессия Аральского моря с соответствующей ей террасой датируется приблизительно от 8 тыс. лет назад (рубеж раннего и среднего голоцена) до 3610±140 – 4956±100 лет назад (Кесь, 1983; Шнитников 1983; Маев, Маева, 1991; Николаев, 1991; и др.), когда существовал сток по Узбою в Каспий. Датировка этой террасы голоценом основана на присутствии в ее осадках раковин Cardium edule L. (вероятно =Cerastoderma isthmicum). По мнению П.В. Федорова (1980), эти отложения в Арале могут быть несколько моложе новокаспийских слоев или равны им по возрасту, а проникновение C. edule в Арал увязывается с расселением его из Каспия, куда он проник от 4–5 до 5–7 тыс. лет назад (Федоров, 1957, 1983; Артамонов, 1976).
По мнению других исследователей, эта терраса более древняя и датирируется поздним плиоценом, возможно апшероном или даже акчагылом (Клейнер, Кравчук, 1966; Кирюхин с соавт., 1966; Кесь, 1969). По данным радиоуглеродного анализа (Пшенин с соавт., 1984), отложения древнеаральской террасы имеют возраст 24820±820 лет.
Анализ наиболее крупных сводок, в которые включены данные по истории видов разных групп организмов, населяющих Арал (Хусаинова, 1957, 1959; Старобогатов, 1970; Шорников, 1973; Мордухай-Болтовской, 1978; Майер, 1979; Иззатуллаев, Старобогатов, 1991; и др.) дает возможность говорить об истоках формирования фауны Арала задолго до возникновения самого Аральского моря. Так, Н.З. Хусаинова (1959) считает, что аральские Dreissena и Hypanis являются реликтами акчагыльского бассейна. По Я.И. Старобогатову (1970), точных датировок появления этой группы в Арале пока нет; в Каспии же они известны с акчагыла. Позднее З. Иззатуллаев и Я.И. Старобогатов (1991) указывают, что Dreissena и Hypanis проникли в Аральскую котловину в апшеронское время и сформировали здесь эндемичные подвиды.
На основе синтеза палеогеографических материалов А.С. Кесь (1969) в жизни Аральского моря выделяет два основных периода – позднеплиоценовый и позднеплейстоценово-современный, указывая, что уровень водоемов этих периодов был неустойчив, и более высоких отметок (до 73 м) достигал позднеплиоценовый бассейн. По Б.И. Пинхасову (1984), в плиоценовое время наиболее обширной являлась апшеронская трансгрессия, достигавшая высоты 73 м над уровнем океана. Судя по последним находкам на территории Казахстана видов моллюсков, обитающих в Аральском море или близких к ним (Старобогатов, Андреева, 1981; Андреева, 1987; Старобогатов, 1994), последним бассейном, в котором продолжала существовать или могла сформироваться фауна, близкая к современной фауне Арала, был позднеплиоценовый бассейн. Так, восточная граница исходного ареала Dreissena polymorpha polymorpha (Старобогатов, Андреева, 1994) в позднем плиоцене (акчагыле) захватывала восточную границу Туранской низменности и западную границу Казахского мелкосопочника. Расселение D. p. polymorpha в восточной части Акчагыльского озера-моря привело к образованию D. balkhashensis Star. et Tolst., найденного недавно в ископаемом состоянии в западном Прибалхашье (Старобогатов, 1994), и части дрейссен Аральского бассейна. Основываясь на морфологическом сходстве D. balkhashensis c D. p. aralensis и D. obtusecarinata, а также и вероятном их филогенетическом родстве, можно предполагать разные пути заселения дрейссенами современного Аральского моря. D. p. aralensis и D. obtusecarinata будучи потомками плиоценовой D. p. polymorpha из восточной части ареала являются, вероятнее всего, восточными вселенцами в Аральское море, в то время как для D. caspia более вероятен западный путь вселения. Считается (Старобогатов, Андреева, 1994), что образование D. caspia на основе одного из подвидов D. polymorpha произошло еще в Меотическом море, в Каспийской котловине вид появился в акчагыльское время, а вселение в Арал датируется не позже раннего плейстоцена с образованием там подвида D. caspia pallasi.
Моллюски р. Hypanis проникли, по З. Иззатуллаеву и Я.И. Старобогатову (1991), в Аральскую котловину в апшероне.
Суждения об истоках и времени проникновения Сerastoderma в Аральское море весьма противоречивы. В научной литературе последних лет появились сведения о времени обитания Сerastoderma в Аральском бассейне, полученные на основании радиоуглеродного анализа ее ракуши. Для центральной котловины Арала слои с Cerastoderma (в оригинале Cardium edule) датируются 4846±90 – 4956±100 лет. В более древних горизонтах с преобладанием пресноводной фауны, предположительный возраст которых 12–14 тыс. лет (максимум верх позднего плейстоцена), Сerastoderma не встречаются (Маев, Маева, 1991).
Наиболее сложен вопрос о времени обитания Сerastoderma в районе древнеаральских террас, датировка которых основывается на присутствии в осадках раковин Сerastoderma. Основываясь, как уже указывалось выше, на положении о вселении Сerastoderma в Арал из Каспия, или синхронизируя обитание Сerastoderma в центральной котловине и на древних террасах, возраст всех осадков, содержащих ракушу Сerastoderma, в том числе и этих террас, определяют не старше среднего голоцена (Рубанов, 1977; Маев с соавт., 1983а; Маев, Маева, 1991; Мамедов, 1991а, б; Аладин, 1996; и др.). Г.Н. Пшениным с соавт. (1984) время обитания Сerastoderma террасы 73 м над уровнем океана радиоуглеродным методом определено в 24820±820 лет, в то время как Б.И. Пинхасов (1984) террасу 73 м над уровнем океана считает соответствующей апшеронской трансгрессии.
В научной литературе существуют две взаимоисключающие точки зрения на истоки и время проникновения Сerastoderma в Арал. Одна из них, основанная на гипотезе Л.С. Берга (1908) о западном пути вселения из Каспия, принимается большинством современных исследователей (Никольский, 1940; Федоров, 1957, 1983; Логвиненко, Старобогатов, 1962, 1964; Артамонов, 1976; Абросов, 1987; Маев, Маева, 1991; и др.).
Вторая точка зрения, вероятно, менее известна широкому кругу исследователей. По гипотезе Н.З. Хусаиновой (1957), Сerastoderma Арала – аборигены Арало-Каспийской области, являются потомками акчагыльских кардиид, вселившимися в Арал во время древнеаральской трансгрессии. Н.З. Хусаинова (1957), основываясь на сообщениях А.Д. Архангельского, Н.П. Луппова, Б.А. Федоровича, высказывает предположение о возможности сохранения этих моллюсков в карстово-суффозионных озерах Сарыкамышской котловины, питавшихся за счет родников, вытекавших из миоценовых отложений, слагавших борта впадин, что подтверждается и более поздними находками А.И. Барановой (1958), однако указывающей западный путь вселения их в Сарыкамыш. Есть также сообщения о находках Cardium edule (вероятно, Cerastoderma isthmicum) в ряде впадин Южного Устюрта (Эберзин, 1959). Узбой же, по мнению Н.З. Хусаиновой (1957), в период своего функционирования служил путем проникновения некоторых «каспийских» форм, в том числе и Cerastoderma, в Каспий из Аральского моря. Некоторые исследователи (Кирюхин с соавт., 1966; Кесь, 1969; Квасов, Трофимец, 1976; и др.) склонны рассматривать Cerastoderma Арала, вслед за Н.З. Хусаиновой (1959), в качестве реликтов акчагыльских форм с допущением более раннего (Квасов, Трофимец, 1976) появления этого моллюска в Арале, чем в Каспии.
Вид Syndosmya segmentum появился в фауне Арала совсем в недавнее время. Моллюск из Бердянских лиманов Азовского моря в количестве 698 тыс. экз. (Карпевич, 1975) вселялся в Аральское море в 1960, 1961 и 1963 гг. [по Е.Л. Марковой (1975) в ? 1959, 1960 и 1962, 1963 гг.]. Выпуск 1960 г. в опресненный залив Джида считается неуспешным, в 1961 и 1963 гг. вселение проводилось в заливе Сары-Чаганак, где моллюск и был обнаружен в 1967 г. (Кортунова, 1970; Атлас …, 1974; и др.). Латентный период при акклиматизации синдосмии был немного короче, чем при акклиматизации этого моллюска в Северном Каспии [вселен в 1947–1948 гг., обнаружен в 1955 г. (Осадчих, 1965)]. Уже в 1967 г. моллюск начинает активно осваивать Малое море (рис. 2.1). Наличие пелагической личинки позволило синдосмии довольно быстро расселиться по дну Аральского моря. Так, в 1968 г. было занято дно залива Сары-Чаганак, в 1971 г. – почти все Малое море и северо-восточная часть Большого моря. В 1972 г. моллюск встречен на большей части акватории, к 1977 г. расселение его в Аральском море завершилось.
<![if !vml]>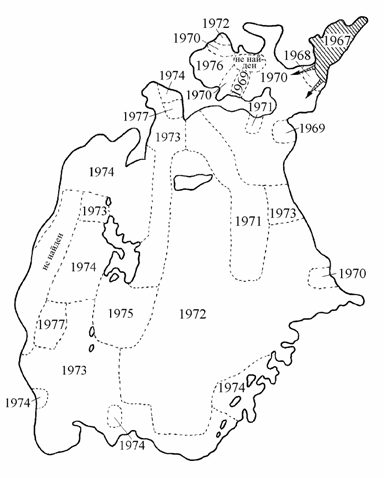 <![endif]>
<![endif]>
Рис. 2.1. Карта-схема расселения Syndosmya segmentum в Аральском море (по: Андреев, Андреева, 1995а). Цифры – годы первого обнаружения вида в данном районе.
Как и в Азовском море, в Арале синдосмия встречалась почти на всех грунтах, наибольшая плотность отмечена также для илистых грунтов. Наиболее благоприятные условия для обитания имелись в районах моря с глубинами 10–20 м, где отмечалась высокая численность и биомасса этого вида (рис. 2.2). Заселению моллюском районов Арала с глубинами более 25 м, вероятно, препятствовало накопление в грунте глубоководной зоны сероводорода, несмотря на отмечаемую способность синдосмии выживать в анаэробных условиях (5–8 дней) и в присутствии сероводорода и аммиака (Воробьев, 1949).
<![if !vml]>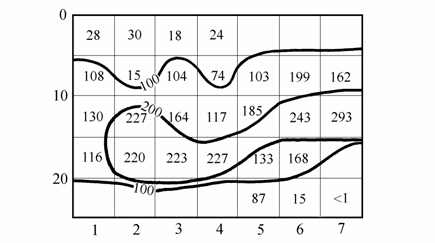 <![endif]>
<![endif]>
Рис. 2.2. Биомасса (г/м2) Syndosmya в зависимости от глубин и грунтов в 1976–1981 гг. (по: Андреев, Андреева, 1997а). По горизонтали — грунты: 1 – коричневый и коричнево-серый илы, 2 – песок, 3 – илистый песок, 4 – песчанистый ил, 5 – серый ил, 6 – темно-серый ил, 7 – черный ил. По вертикали — глубина, м.
Успешному освоению акватории Арала способствовали также такие свойства моллюска, как эвригалинность и эврибионтность. Популяция синдосмии из литоральной зоны южной Бретани обитает при температуре воды 3.5–20° и солености 25–36 ‰ (Denis, 1981). В Аральском море моллюск был встречен в осолоненных районах по всему исследованному диапазону солености от 10 до 40 ‰, температура в местах обитания в летний период времени колебалась от 5.7 до 28.0° (Андреев, Андреева, 1995а). Как и при расселении нереиса (Андреев, Андреева, 1992), здесь определенное влияние на скорость распространения оказывали гидрологические условия (особенно сила и направление течений), слагавшиеся в период нереста и пребывания в планктоне пелагической личинки, однако большое значение имело и наличие подходящего субстрата для оседания личинок.
Итак, подводя итог небольшому экскурсу в прошлое Аральского моря, следует отметить, что и по последним публикациям можно представить только в общем историю этого водоема, а также пути формирования его фауны и время проникновения отдельных элементов. Остается дискуссионным вопрос о времени и водоеме, из которого проникло большинство двустворчатых моллюсков в Аральское море. В научной литературе последних лет преобладает западная гипотеза вселения двустворок в Арал из Каспия (Иззатуллаев, Старобогатов, 1991; Маев, Маева, 1991; Федоров, 1980, 1983; Мамедов, 1991а, б; и др.).