
Проблемы Эволюции |
О взрывном видообразовании, протекающем на наших глазах в высыхающем Аральском море. Омск: Изд-во Омского государственного педагогического университета, 2003. 382 с.
Об обитании моллюсков рода Hypanis (=Adacna) в Аральском море на начало двадцатого века имеются весьма противоречивые сведения, суммируя которые можно отметить, что систематиками конца девятнадцатого – начала двадцатого века для Аральского моря указывались Adacna minima Ostr., Adacna vitrea Eichw., Adacna laeviuscula Eichw. (Basiner, 1848; Мартенс, 1874; Аленицин, 1876; Остроумов, 1907; Берг, 1908).
После исследований коллекций Аральской Научно-Промысловой экспедиции С.А. Сидоров (1929) для Аральского моря описывает две разновидности Adacna minima Ostr., обитающих в соленой воде Арала и в сильно опресненных его участках, а также указывает Adacna vitrea Eichw.
В.И Жадин (1952) для Аральского моря приводит обитание при солености 5–15 ‰ Adacna vitrea Eichw. и Adacna laeviuscula Eichw.
Н.З. Хусаинова (1958) отмечает трудности выяснения систематической принадлежности аральских Adacna из-за большого разнообразия в морфологии раковин. Большинство особей по ее мнению подходило под описание Adacna minima Ostr., изредка встречались особи очень сходные с Adacna vitrea Eichw., а некоторые экземпляры по форме синуса напоминали Adacna laeviuscula Eichw. Каспийского моря.
Я.И. Старобогатовым (1974) для Аральского моря определено два вида Hypanis из подрода Adacna, один из которых распадается на два подвида: H. vitrea bergi Star., H. minima sidorovi Star., H. m. minima (Ostr.), при этом указано, что H. m. minima распространен по всему Аралу на мягких грунтах на глубинах свыше 10–12 м, а H. m. sidorovi обитает в прибрежных районах на глубинах до 10 м.
Анализ сохранившихся сборов за май 1965 г. и собственных за 1976–1980 гг. подтверждает наличие в Аральском море трех форм моллюсков рода Hypanis (рис. 5.1). Совместное обитание в водоеме двух подвидов одного вида с пелагической личиночной стадией развития и, тем более, их совместное нахождение в одной дночерпательной пробе на станции 31 в мае 1965 г. в свете современной теории видообразования ставит под сомнение таксономический статус этих форм. Предлагается поднять до видового ранга статус H. minima sidorovi и именовать его Hypanis sidorovi Star., тем более , что все три аральские формы достаточно хорошо различаются формой лигамента (рис. 5.2), считающегося надежным систематическим признаком (Petersen, 1958; Михайлова, 1985).
Hypanis vitrea bergi Star., 1974 (рис. 5.1, 1–2)
Adacna minima Ostr.: Sidorov, 1929 (частью); Hypanis vitrea bergi Starobogatov, 1974.
<![if !vml]>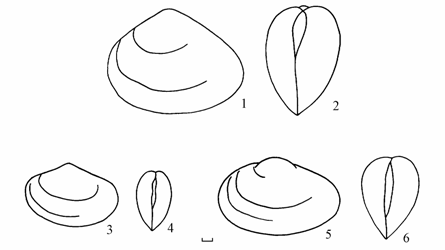 <![endif]>
<![endif]>
Рис. 5.1. Hypanis Аральского моря.
1–2 — H. vitrea bergi, 3–4 — H. minima minima, 5–6 — H. sidorovi. 1, 3, 5 – вид сбоку, 2, 4, 6 – вид сзади. Масштаб 1 мм.
По Я.И. Старобогатову (1974: 245): «Раковина розовая или белая, просвечивающая, тонкостенная, овальная, почти равносторонняя, довольно высокая (высота составляет 0.78–0.85 длины), плоская (выпуклость составляет 0.40–0.50 длины и 0.48–0.60 высоты). Поверхность покрыта частыми, плоскими, сильно сглаженными ребрами. Раковина зияет сзади и спереди. Синус глубокий, доходящий до границы задней трети длины раковины.
<![if !vml]>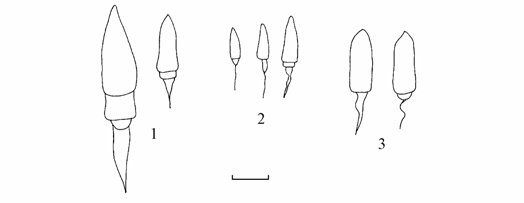 <![endif]>
<![endif]>
Рис. 5.2. Лигаменты Hypanis Аральского моря.
1 – H. vitrea bergi, 2 – H. minima minima, 3 – H. sidorovi. Масштаб 1 мм.
Длина до 20 мм
Подвид очень похож на северокаспийский Н. v. glabra (Ostr.), от которого отличается более частыми и сглаженными ребрами и несколько более широким синусом. Известен по единичным находкам живых особей в районе дельты Амударьи и пустых раковин на западном берегу Арала, типовое местонахождение: район колодца Кара-Кибир.»
В 1976–1978 гг. H. v. bergi единично встречался в северо-западном районе глубоководной впадины на черном иле и вдоль восточного побережья на песчанистом грунте на глубинах 12–31 м. С 1980 г. в сборах не отмечался.
Hypanis minima minima (Ostr., 1907) (рис. 5.1, 3–4)
Adacna minima Ostroumoff, 1907; Adacna minima Ostr.: Sidorov, 1929 (частью); Hypanis minima minima (Ostr.): Starobogatov, 1974.
По Я.И. Старобогатову (1974: 246): «Раковина белая, просвечивающая, очень тонкая, удлиненно-овальная (высота составляет 0.67–0.74 длины), мало выпуклая (выпуклость составляет 0.40–0.50 длины и 0.60–0.70 высоты), однако при сопоставлении боковых контуров более выпуклая, чем Н. vitrea, сзади зияющая с узкой мало выступающей слегка сдвинутой вперед макушкой. Ребра незаметны, вместо них видны лишь тонкие, радиально расходящиеся от макушки штрихи. Синус широкий и довольно глубокий, доходящий до границы задней трети длины раковины. Рудименты кардинального зуба в правой створке сохраняются.
Длина до 15 мм».
H. m. minima найден в мае 1965 г. на глубине 21.8 и 26 м, на песчанистом и черном иле (ст. 42 и ст. 14 соответственно), также на станциях 20 и 31; в июле 1977 г. – в районе полуострова Куланды на глубине 14.3 м на песчаном грунте при солености 14.4 ‰. В последующем не встречен.
Hypanis sidorovi Star., 1974 (рис. 5.1, 5–6)
Adacna laeviuscula Martens, 1874; Hypanis minima sidorovi Starobogatov, 1974.
По Я.И. Старобогатову (1974: 246): «Раковина белая, слабо просвечивающая, довольно тонкостенная, удлиненно-овальная (высота составляет 0.68–0.77 длины), мало выпуклая (выпуклость составляет 0.40–0.50 длины и 0.60–0.70 высоты), сзади зияющая, с довольно широкой заметно сдвинутой вперед макушкой. Ребра сильно сглаженные, но заметные. Синус широкий и довольно глубокий, доходящий до границы задней трети длины раковины. Рудименты кардинального зуба в правой створке сохраняются.
Длина до 20 мм.»
H. sidorovi найден в мае 1965 г. на станциях 35 и 31; в 1976–1977 г. — в северо-западном районе глубоководной впадины в заливе Тщебас и вдоль восточного побережья на песчаном грунте на глубине 12–31 м. В последующие годы не обнаружен.
Итак, в 1970-е годы в Аральском море обитало три вида моллюсков рода Hypanis: H. vitrea bergi, H. minima minima, H. sidorovi; один из которых был эндемичен, а два другие сформировали здесь эндемичные подвиды.
Для моллюсков рода гипанис в Аральском море, по наблюдениям Е.А. Яблонской (1960), были благоприятны грунты на всех глубинах открытой части моря за исключением биотопов с возможным дефицитом кислорода. В середине 1950-х годов их доля в общей биомассе зообентоса составляла 17.7–31.7 %. Гипанисы были распространены довольно равномерно по всей акватории моря, лишь в северной части наблюдались скопления с биомассой 20 г/м2 и более. Однако уже в 1964–1965 гг. (рис. 5.3) наряду с понижением биомассы этих моллюсков в среднем по водоему наблюдалось сокращение и их ареала, особенно резкое в восточной и западной частях моря. В 1966–1967 гг. распространение гипанисов было в виде отдельных пятен, биомасса снизилась с 8.2 г/м2 в 1954 г. до 0.2 г/м2 в 1967 г. (см. табл. 3.2). В 1968–1975 гг. наблюдалось небольшое расширение ареала и увеличение средней биомассы гипанисов. В последующие годы, когда средняя соленость воды в море стала приближаться к сублетальной для этой группы видов, численность и биомасса аральских гипанисов быстро снизилась и в 1980 г. в период стандартной съемки не было найдено ни одного экземпляра.
Результатом критической ситуации в донных сообществах Арала, обусловленной массовым размножением в 1960-е годы в прибрежье сорных рыб-вселенцев, явилось резкое снижение с середины 1960-х годов частоты встречаемости и биомассы всех видов Hypanis. Восстановление в 1968 г. ареала этих моллюсков вновь сменяется его значительным сокращением уже в следующем году (см. рис. 5.3). В последующем наблюдаются флуктуационные колебания зон обитания Hypanis, обусловленные, вероятно, несколько различной солеустойчивостью отдельных видов и неодновременным достижением различных районов моря пороговой для этих видов солености. Наличие пелагической личинки при дестабилизации среды обитания приводит к обитанию видов вне пределов свойственных им зон. Начиная с 1976 г. в сборах по стандартной сетке станций эти моллюски присутствуют лишь единично, постепенно выпадая из фауны Арала. Последним вымирает, судя по находкам, H. v. bergi.
<![if !vml]>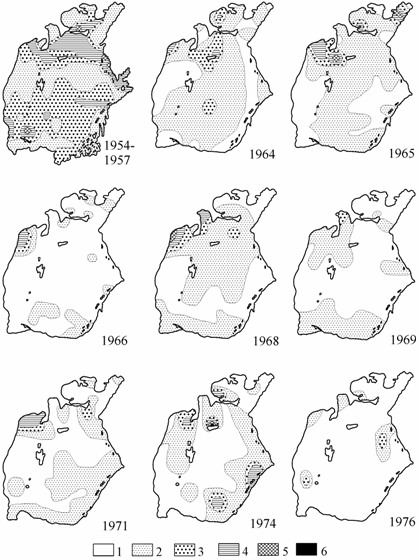 <![endif]>
<![endif]>
Рис. 5.3. Распределение биомассы Hypanis в Аральском море
летом (по: Андреева, 1989; 1954–1957 гг. из: Яблонская, 1960).
1 — отсутствует, 2 — 0.1–5, 3 — 5.1–10, 4 — 10.1–20, 5 — 20.1–40, 6 — более 40 г/м2.
В силу своего формирования в пределах олигогалинной водной массы, не обладая физиологическими механизмами, обеспечивающими обитание, а главное размножение, при более высоких соленостях, эти моллюски оказались не способными преодолеть барьер критической солености 12–14 ‰. По имеющимся коллекциям невозможно проследить влияние приближения критической солености на отдельные виды в плане каких-либо морфологических перестроек или возрастания изменчивости. Хотя эти процессы, вероятнее всего, имели место при осолонении моря. Можно отметить лишь наличие уже в 1965 г. наружных дефектов створок почти у всех особей H. m. minima, а собранные 9 особей этого подвида на станции 42 имели очень уродливые створки с искривленными макушками.